Кінцевий мозок ссавців. Головний мозок ссавців Передній мозок ссавців досяг високого розвитку
Розвинена нервова система - одна з основних прогресивних ознак ссавців, завдяки яким вони займають вищу позицію в тваринному світі.
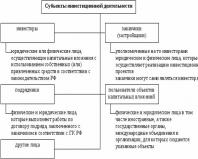
Центральна нервова система
Нервова система ссавців поділяється на центральну та периферичну.
Головний та спинний мозок утворюють центральну нервову систему (ЦНС).
Головний мозок
Головний мозок має ті ж 5 відділів, що і мозок інших хребетних:
- передній;
- проміжний;
- середній;
- мозок;
- довгастий.

Мал. 1. Головний мозок ссавця.
Але, якщо у рептилій маса спинного мозку приблизно дорівнює масі головного, то у ссавців головний мозок важить у 3 – 15 разів більше.
ТОП-3 статтіякі читають разом з цією
Збільшення маси та обсягу головного мозку ссавців відбувається за рахунок збільшення кори великих півкуль переднього мозку.
У вищих загонів (хижі, примати, ластоногі та китоподібні) кора утворює борозни, які значно збільшують площу її поверхні.
Попереду півкуль знаходяться нюхові частки. У ссавців вони найбільше розвинені, що пов'язано з великим значенням у них нюху та язика запахів.
Відношення маси півкуль до маси всього головного мозку у звірів по-різному:
- їжаки 48%;
- білки 53%;
- вовки 70%;
- дельфіни 75%.
Мозок також значно розвиненіший, ніж в інших класів. Він має кілька відділів та складчасту кору. У мозочку відбувається управління складними рухами.
Слово кора щодо структур мозку має умовне значення. Вона не є захисним шаром. У ній знаходяться центри, що забезпечують складну поведінку ссавців.
У проміжному мозку знаходяться:
- гіпофіз;
- епіфіз;
- гіпоталамус.
Вони регулюють:
- зростання організму;
- обмін речовин;
- тепловіддачу;
- сталість внутрішнього середовища організму.
Середній мозок розділений борознами на 4 пагорби. У ньому розташовані центри слуху та зору.
Довгастий мозок є продовженням спинного та керує диханням, травленням та кровообігом.
Спинний мозок
Спинний мозок знаходиться у каналі хребта. Він виконує провідну та рефлекторну функції.

Мал. 2. Спинний мозок ссавця.
Спинний мозок передає імпульси від мозку до органів і назад.
Рефлексами спинного мозку називають прості реакції на подразнення, наприклад, чухання у відповідь на почуття сверблячки.
Периферична нервова система (ПНР)
Усі нерви, нервові закінчення та нервові вузли, що лежать за межами ЦНС, відносять до периферичної нервової системи.

Мал. 3. Схема периферичної нервової системи ссавців.
До неї відносять нерви, що відходять від головного (12 пар) та спинного (31 пара) мозку.
Ці нерви трьох типів:
- чутливі;
- рухові;
- змішані.
Так, нюховий, слуховий та зоровий нерви є чутливими. Окоруховий нерв – руховим. Під'язичний та язикоглотковий нерви – змішані.
Що ми дізналися?
Ми розповіли коротко про нервову систему ссавців. Вона відрізняється значним розвитком всіх відділів головного мозку, особливо мозочка та переднього мозку.
Тест на тему
Оцінка доповіді
Середня оцінка: 4.7. Усього отримано оцінок: 117.
Продовгуватий мозок. Як і у представників нижчестоящих класів, від дна проміжного мозку відходять зорові нерви, що утворюють перехрест, і за ними знаходиться лійка, до якої прикріплюється гіпофіз, тоді як над проміжним мозком розташовується на довгій ніжці епіфіз.
Порожнина проміжного мозку, або третій шлуночок, з обох боків має потужні скупчення мозкової речовини, що мають назву зорових горбів (thalami optici). Таким чином, проміжний мозок має подібну будову з відповідним мозком плазунів та птахів.
Середній мозок, Навпаки, відрізняється порівняно дуже невеликими розмірами і дах його, крім поздовжньої борозни, має ще поперечну борозну. Завдяки цьому у кролика, як і у всіх ссавців, замість двоолмію, властивого представникам інших класів, дах середнього мозку представлений чотиригір'ям (corpus quadrigeminum). Передні пагорби несуть зорову функцію, а задні - слухову. Порожнина середнього мозку, або сильвієвий водопровід, є лише вузькою щілиною.
Мозжечокскладається з середньої непарної частини - черв'ячка - і двох бічних частин, які дуже великі і позначаються як півкуля мозочка (hemisphaerae cerebelli). Від них відходять убік бічні придатки (flocculi).
Продовгуватий мозоквідрізняється від такого у представників нижчестоящих класів тим, що з боків четвертого шлуночка окремоляються поздовжні пучки нервових волокон, що йдуть до мозочка і носять назву задніх ніжок мозочка (crura medullo-cerebellaria), на нижній поверхні довгастого мозку відокремлюються парні поздовжні валики — піраміди (pyramis), а попереду них лежить поперечне піднесення, що складається пов'язують під довгастим мозком праву і ліву півкулі мозочка. Піднесення це характерно для ссавців і зветься еаролієвим мостом (pons varolii).

. I-згори; II - знизу; III - збоку; IV - поздовжній розріз (за Паркером):
1 - великі півкулі, 2 - нюхові частки, 3 - зоровий нерв, 4 - епіфіз, 5 - середній мозок-четверогормие, 6 - мозок, 7 - довгастий мозок, S - гіпофіз, 9 - варолієв міст, 10 - мозкова лійка, 11 - мозолисте тіло, 12 - зорові горби
Головні нерви. У кролика вже 12 пар головних нервів, так як XI пара - додатковий нерв (nervus accessorius), не цілком диференційований у птахів і плазунів, отримує у ссавців повнерозвиток. Він відходить від боків довгастого мозку приблизно на рівні XII пари. Інші головні нерви мають типові місця відходження.
Органи відчуттів. Для кролика, як і для гризунів, характерний сильний розвиток дотикового волосся - вібріс - на голові у вигляді так званих вусів, на верхній і нижній губах, підборідді, на щоках і бровах. Серед органів чуття, як і в більшості ссавців, провідну роль грають органи нюху; у нюхової порожнини, як сказано вище, є складний лабіринт нюхових раковин. Високого ступеня досконалості досягають також органи слуху, що мають складно звивистий равлик, звукопровідний апарат в середньому вусі з трьох слухових кісточок, кісткові слухові барабани і великі рухливі зовнішні вуха.
Ще цікаві статті
Головний мозок ссавців, зберігаючи загальні хребетним тваринам риси, відрізняється важливими особливостями, які змушують виділяти їх у особливий кортикальний тип.
У головному мозку ссавців найбільшої величини та складності досягає передній мозок, у якому більшість мозкової речовини зосереджена в корі півкуль, тоді як смугасті тіла відносно невеликі. Дах (кора) переднього мозку формується шляхом розростання нервової речовини стінок бічних шлуночків. Мозковий звід, що утворюється таким чином, носить назву вторинного склепіння або неопаліуму (neopallium); його зачатки з'являються у амфібій і більш помітні у рептилій та птахів. Він складається з нервових клітин та безм'якотних волокон (сіра речовина мозку). Обидві півкулі пов'язані між собою комісурою з білих (міелінізованих) волокон, званої мозолистим тілом. Тіла нейронів у корі півкуль розташовані пошарово, утворюючи своєрідні "екранні структури" (рис. 106). Така організація мозку дозволяє просторово відображати зовнішній світ на основі інформації, що надходить від органів чуття. Екранні структури характерні найважливіших мозкових центрів ссавців, тоді як в інших хребетних вони зустрічаються рідше, переважно у зорових центрах. Нова кора великих півкуль служить центром вищої нервової діяльності, що координує роботу інших відділів мозку (рис. 107). Лобові частки здійснюють управління спілкуванням тварин, зокрема акустичним; у людини вони пов'язані з промовою, тобто другою сигнальною системою.
Кора майже всіх ссавців утворює більшу чи меншу кількість борозен, що збільшують її поверхню. У найпростіших випадках є одна сильвієва борозна, що відокремлює лобову частку від скроневої; потім з'являється роландова борозна, що розділяє лобну і потиличну частки, та ін У приматів і зубастих китів число борозен особливо велике. Неопаліум ссавців більшою мірою, ніж комплекс кори середнього та переднього мозку птахів, забезпечує вищу нервову діяльність, накопичуючи сліди одиничних збуджень та їх поєднань, тобто збагачуючи так звану оперативну пам'ять. Це відкриває можливість на її основі обирати за нової ситуації оптимальні рішення. Найчастіше вони становлять нові комбінації вже відомих елементів. Виникнення у ссавців вищих асоціативних центрів - нової кори - не вело до ліквідації центрів, які керують інстинктивними актами, але лише підкорило їх вищому контролю.
Інші частини переднього мозку мають відносно менші розміри, але зберігають своє значення. Нюхові частки розташовані в передній частині внизу (нюхові цибулини та передні базальні ядра). Смугасті тіла ссавців виконують функцію регуляції інстинктивних реакцій під контролем кори півкуль.
Проміжний мозок невеликий і зверху прикритий півкулями переднього мозку. У ньому розташовані третій шлуночок і зорові горби, через які проходить зоровий тракт і де відбувається первинна обробка зорової інформації. У даху розташований невеликий епіфіз (секреторний орган). У дні проміжного мозку (гіпоталамус) розташовані вегетативні центри, що беруть участь у регуляції процесів метаболізму та терморегуляції. Там же знаходиться лійка, тісно поєднана з ендокринною залозою – гіпофізом. Останній бере участь у сезонній перебудові метаболізму та регуляції періодичних явищ (линьки, сплячки, розмноження, міграцій). Секретування ним відповідних гормонів відбувається під впливом речовин, що виділяються нейросекреторними клітинами гіпоталамуса і що надходять у гіпофіз кровоносних судин, що зв'язує його з гіпоталамусом.
Середній мозок малий; його дах поперечними борознами поділено на четверохолміе , в якому передні горби утворюють слабо виражену зорову кору, задні служать слуховими центрами, підлеглими контролю переднього мозку (рис. 108).
Мозок великий і складається з серединного черв'ячка і прилеглих до нього парних півкуль з бічними придатками. Як і в інших хребетних, з мозком пов'язана підтримка м'язового тонусу, пози, рівноваги та пропорційності рухів частин тіла. Тісні зв'язки мозочка з корою півкуль свідчать про існування вищого контролю і тут.
Довгастий мозок дає початок більшості черепних нервів (V – XII). У ньому розташовані центри дихання, роботи серця, травлення і т. п. З боків порожнини четвертого шлуночка відокремлюються пучки нервових волокон, що йдуть до мозочка і утворюють його задні ніжки. Нервові тракти пов'язують довгастий мозок зі спинним мозком.
Як і в інших хребетних, відносні розміри головного мозку збільшуються при зменшенні розмірів тіла та зростанні напруженості терморегуляції (Стрільників). Так, у великих комахоїдних, маса головного мозку становить близько 0,6% маси тіла, а у дрібних – до 1,2, у великих китоподібних – близько 0,3, а у дрібних – до 1,7% і т.д. Маса мозку приматів становить 0,6-1,9% від маси тіла, а в людини – близько 3%. У всіх ссавців маса переднього мозку перевищує масу інших відділів головного мозку: у різних групах вона становить 52-72% загальної маси мозку головного; у приматів цей показник зростає до 76-80%, а у людини-до 86% (Нікітенко, 1969).
Співвідношення мас головного та спинного мозку максимально у людини (45: 1), високо у приматів та китоподібних (10-15: 1) і нижче у хижаків, комахоїдних (3-5: 1) та копитних (2,5: 1). У рептилій воно завжди менше одиниці, а у птахів становить 1:2 - 5:1. Спинний мозок за допомогою провідних шляхів (біла речовина) пов'язаний з руховим центром кори півкуль, що здійснює вищий контроль над руховими актами та керування складними рухами. Спинні стовпи білої речовини складаються з волокон, що несуть імпульси від органів чуття і ентеро-рецепторів (аферентна інформація), тоді як у черевних стовпах переважають волокна, що несуть імпульси від мозку до м'язів та інших виконавчих органів (еферентна інформація). Короткі провідні шляхи пов'язують сусідні сегменти. Контроль вищих центрів мозку над роботою спинного мозку досягає у ссавців найбільшого рівня.
Ссавці мають 12 пар головних черепних нервів; розвивається XI пара - додаткові нерви (п. accessorius). Крім іннервації основних органів чуття (нюху, зору, слуху) і м'язової системи головні нерви беруть участь в утворенні вегетативної нервової системи, що контролює так звані вегетативні процеси, що не підкоряються вольовому (довільному) контролю. Парасимпатична нервова система утворена черепними нервами довгастого мозку та спинномозковими нервами крижового відділу.
 Загальні характеристики: Мозок ссавців можна поділити на два типи: спинний та головний мозок. У свою чергу головний мозок поділяється на: 1. Довгастий мозок 2. Задній мозок 3. Середній мозок 4. Проміжний мозок 5. Кінцевий мозок
Загальні характеристики: Мозок ссавців можна поділити на два типи: спинний та головний мозок. У свою чергу головний мозок поділяється на: 1. Довгастий мозок 2. Задній мозок 3. Середній мозок 4. Проміжний мозок 5. Кінцевий мозок
 Мозок кролика: I – зверху; II – знизу; III – збоку; IV – поздовжній розріз. 1 – великі півкулі; 2 - нюхові частки; 3 - зоровий нерв; 4 – епіфіз; 5 – середній мозок; 6 - мозок; 7 - довгастий мозок; 8 – гіпофіз; 9 - вароліїв міст; 10 - мозкова лійка; 11 - мозолисте тіло
Мозок кролика: I – зверху; II – знизу; III – збоку; IV – поздовжній розріз. 1 – великі півкулі; 2 - нюхові частки; 3 - зоровий нерв; 4 – епіфіз; 5 – середній мозок; 6 - мозок; 7 - довгастий мозок; 8 – гіпофіз; 9 - вароліїв міст; 10 - мозкова лійка; 11 - мозолисте тіло

 Мозковий стовбур складається з довгастого мозку, варолієвого мосту та середнього мозку. Часто в нього включають мозок.
Мозковий стовбур складається з довгастого мозку, варолієвого мосту та середнього мозку. Часто в нього включають мозок.
 Мозок У ссавців мозок складається з черв'яка (середня частина) і двох бічних часток. У однопрохідних середня частина більше бічних, у сумчастих вони приблизно подібні, а відмінністю вищих ссавців є збільшення бічних часток, пов'язаних з корою великих півкуль. Для збільшення площі поверхні мозочка, як і великі півкулі, покритий борознами та звивинами. Функції мозочка: Хробак відповідає позу, тонус осьових м'язів тіла та проксимальних кінців м'язів Координація рухів Регуляція рівноваги
Мозок У ссавців мозок складається з черв'яка (середня частина) і двох бічних часток. У однопрохідних середня частина більше бічних, у сумчастих вони приблизно подібні, а відмінністю вищих ссавців є збільшення бічних часток, пов'язаних з корою великих півкуль. Для збільшення площі поверхні мозочка, як і великі півкулі, покритий борознами та звивинами. Функції мозочка: Хробак відповідає позу, тонус осьових м'язів тіла та проксимальних кінців м'язів Координація рухів Регуляція рівноваги

 Середній мозок входить до складу ствола головного мозку. Є зоровим центром мозку. Вентральну частину складають масивні ніжки мозку, основну частину яких займають пірамідні шляхи. Між ніжками знаходиться міжніжкова ямка (лат. fossa interpeduncularis), з якої виходить III нерв. Дорсальна частина - платівка чотирипагорби, дві пари горбків. Верхні пагорби зорові, вони більші за нижні (слухові). З дорсального боку межі з мостом відходить IV нерв. У глибині покришки середнього мозку (під четверохолмієм) знаходяться ядра окорухових нервів, червоні ядра (управління рухом), чорна речовина (ініціація рухів), ретикулярна формація.
Середній мозок входить до складу ствола головного мозку. Є зоровим центром мозку. Вентральну частину складають масивні ніжки мозку, основну частину яких займають пірамідні шляхи. Між ніжками знаходиться міжніжкова ямка (лат. fossa interpeduncularis), з якої виходить III нерв. Дорсальна частина - платівка чотирипагорби, дві пари горбків. Верхні пагорби зорові, вони більші за нижні (слухові). З дорсального боку межі з мостом відходить IV нерв. У глибині покришки середнього мозку (під четверохолмієм) знаходяться ядра окорухових нервів, червоні ядра (управління рухом), чорна речовина (ініціація рухів), ретикулярна формація.
 Проміжний мозок Складається з: Таламічний мозок 1. Таламус (зоровий бугор) 2. Епіталамуса 3. Метаталамус Гіпоталамус Третій шлуночок
Проміжний мозок Складається з: Таламічний мозок 1. Таламус (зоровий бугор) 2. Епіталамуса 3. Метаталамус Гіпоталамус Третій шлуночок
 Таламус – складається із сірої речовини, парне утворення яйцеподібної форми. Центр чутливості. Епіталамус - утворює шишкоподібне тіло (епіфіз), заліза внутрішньої секреції, відповідає за синхронізацію біоритмів організму. Метаталамус – утворений латеральним та медіальним колінчастими тілами, центр слуху. Гіпоталамус - включає соскоподібні тіла, що є підкірковими центрами нюху, гіпофіз, зоровий перехрест, II пари черепних нервів, сірий бугор, що являє собою вегетативний центр обміну речовин і терморегуляції. У гіпоталамусі містяться ядра, що контролюють ендокринні та вегетативні процеси. Третій шлуночок – порожнина проміжного мозку. Він є вузьким, розташованим в сагітальній площині щілиноподібним простором. Третій шлуночок має п'ять стінок.
Таламус – складається із сірої речовини, парне утворення яйцеподібної форми. Центр чутливості. Епіталамус - утворює шишкоподібне тіло (епіфіз), заліза внутрішньої секреції, відповідає за синхронізацію біоритмів організму. Метаталамус – утворений латеральним та медіальним колінчастими тілами, центр слуху. Гіпоталамус - включає соскоподібні тіла, що є підкірковими центрами нюху, гіпофіз, зоровий перехрест, II пари черепних нервів, сірий бугор, що являє собою вегетативний центр обміну речовин і терморегуляції. У гіпоталамусі містяться ядра, що контролюють ендокринні та вегетативні процеси. Третій шлуночок – порожнина проміжного мозку. Він є вузьким, розташованим в сагітальній площині щілиноподібним простором. Третій шлуночок має п'ять стінок.

 1 – кора 2 – зовнішній шар кори 3 – лімбічна система 4 – таламус 5 – гіпоталамус 6 – четверогір'я 7 – мозочок 8 – довгастий мозок
1 – кора 2 – зовнішній шар кори 3 – лімбічна система 4 – таламус 5 – гіпоталамус 6 – четверогір'я 7 – мозочок 8 – довгастий мозок

 Кінцевий мозок складається з двох півкуль великого мозку (покритих корою), мозолистого тіла, смугастого тіла та нюхового мозку. Кора великих півкуль розподіляється на давню, стару та нову кору. Стародавня кора виконує найнеобхідніші функції, такі як розкриття очей. Стара кора відповідає за вияв емоцій. Нова кора відповідає за найвищу нервову діяльність. Мозолисте тіло з'єднує дві півкулі мозку. Відсутня у сумчастих та клоочних. Смугасте тіло виконує функції гіпертонусу скелетних м'язів, порушення складних рухових реакцій та харчовидобувної поведінки, гальмування формування умовних рефлексів. Нюховий мозок відповідає за всі структури кінцевого мозку, пов'язані з нюхом.
Кінцевий мозок складається з двох півкуль великого мозку (покритих корою), мозолистого тіла, смугастого тіла та нюхового мозку. Кора великих півкуль розподіляється на давню, стару та нову кору. Стародавня кора виконує найнеобхідніші функції, такі як розкриття очей. Стара кора відповідає за вияв емоцій. Нова кора відповідає за найвищу нервову діяльність. Мозолисте тіло з'єднує дві півкулі мозку. Відсутня у сумчастих та клоочних. Смугасте тіло виконує функції гіпертонусу скелетних м'язів, порушення складних рухових реакцій та харчовидобувної поведінки, гальмування формування умовних рефлексів. Нюховий мозок відповідає за всі структури кінцевого мозку, пов'язані з нюхом.

Кінцевий мозок ссавців має найскладніші кіркові формації півкуль. Основу цитоархітектонічного поділу кори півкуль розробив на початку XX ст. К. Бродман. Він розділив кору на дві основні області: гомогенетичну, для якої характерно шість - семишаровий будову, і гетерогенетичну, де така будова відсутня. Гомогенетична кора поділяється на гомотипічну (шестишарову будову зберігається і в дорослому стані) та гетеротипічну (у процесі ембріогенезу кількість шарів змінюється). Кожна з цих територій поділяється на області, а області на поля, що розрізняються за будовою, зв'язками, функціональною значимістю. Так, у приматів виділено 11 областей, що включають 52 поля.
За класифікацією І. Н. Філімонова основна область кори, позначена ним як нова кора (неокортекс, ізокортекс), відповідає гомогенетичній корі за Бродманом. Області старої (архікортекс) і стародавньої (палеокортекс) кори гомологічні лише частини гетерогенетичної кори. Решту кори займає так звана проміжна кора (мезокортекс), що складається з періархікортексу, що відокремлює нову кору від старої, і перипалеокортексу, що відокремлює нову кору від стародавньої.
До складу нової кори, на думку І. Н. Філімонова, має бути включений весь паліум, у межах якого стінка мозку вже
на ранніх етапах розвитку характеризується наявністю основних верств Гисса (матрикс, проміжний шар, коркова пластинка, крайової шар). Стара і давня кора, на відміну нової, на ранніх стадіях розвитку характеризується неповнотою будови стінки мозку. Межкова кора має перехідний тип будови.
До палеокортексу відносяться піриформна кора і препіриформна область півкулі, а також нюхова цибулина, переднє нюхове ядро, нюховий горбок, септум, частина амігдали і ряд дрібних ядер вентромедіальної стінки півкулі. До складу архікортексу у ссавців включають райони гіпокампу, зубчастої фасції, субикулуму. Періархікортекс представлений пре-і парасубікулумом, taenia tecta, енторинальною та цингулярною корою; перипалеокортекс – перехідною інсулярною областю. Дослідження великої кількості ссавців показало, що розподіл кори на зазначені вище основні області всюди зберігає своє значення і проблема їхньої гомології у представників різних груп ссавців не зустрічає суттєвих труднощів.
У 70-ті роки, розвиваючи ідеї І. Н. Філімонова та інших дослідників, швейцарський нейроморфолог Ф. Занідес сформулював теорію чотирьох послідовних стадій розвитку кори півкуль ссавців. Відповідно до цієї теорії неокортекс має складне походження і його формації розвиваються з двох зачатків: архікортексу та палеокортексу. Архікортикальна «половина» - попередник неокортикальних формацій медіальної стінки півкулі, палеокортикальна «половина» - вентральної та латеральної. Перша стадія на шляху формування неокортексу («ізокортексу» в термінології Ф. Занідеса) – утворення двошарової проміжної кори (мезокортексу), що складається з періаллокортексу – медіально, перипалеокортексу – латерально (рис. 110). Потім утворюються формації вже багатошарової кори, представлені в дорослому мозку структурами цингулярної кори – медіально та інсулярної (острівцевої) – латерально. Далі на їх основі формуються власне відділи шестишарового ізокортексу. Він включає дві зони: перехідну (произокортекс, прокониокортекс), що включає парацингулярную (лімбічну) і параинсулярную (інсулярну) кору, і основну зону, представлену структурами справжньої нової кори - коніокортексу. На останньому етапі формування нової кори зі структур перехідних областей формуються первинні та вторинні сенсорні кіркові зони. У межах розвиваються й інші поля і зони нової кори. Таким чином, нова кора ссавців має подвійне походження, і кожна її ділянка має певні структурно-функціональні особливості в залежності від видалення первинних зон формування даної області кори.
Нова кора (Neocortex). В еволюції ссавців нова кора завдяки своєму інтенсивному зростанню відтіснила стару та давню кору на медіальну та вентральну поверхні півкулі. В результаті сформувалися дві основні борозни: гіпокампова (sh), що відокремлює стару кору від нової, і ринальна (srh), що відокремлює давню кору від нової. Ці борозни чітко виражені у кінцевому мозку всіх ссавців. Далі відбувалося утворення борозен та звивин у межах кіркових областей. За ступенем розвитку борозен і звивин півкулі мозку ссавців поділяються на два типи: лісенцефалічний, для якого характерна відсутність або дуже слабкий розвиток борозен на поверхні півкулі, і гіренцефалічний - з більш менш розвиненою системою борозен і звивин. Перший тип характерний для однопрохідних, нижчих сумчастих, комахоїдних, рукокрилих, гризунів. Інші ссавці мають на поверхні півкуль систему борозен і звивин. Вивчення конфігурації звивин і борозен мозку різних ссавців має важливе значення, проте спроби стандартизувати номенклатуру борозен і звивин поки що успішні лише щодо мозку людини та кількох видів приматів (рис. 111).

Мал. Ill
Малюнок основних борозен (Sulcus)і звивин (gyrus)на бічній (Л) та медіальній (Б) поверхнях головного мозку приматів (людина. Нігтю supiens)
Щодо інших ссавців проблема номенклатури борозен та звивин дуже актуальна. Є кілька причин невідповідності номенклатур: 1 - види, що вивчаються (навіть близькоспоріднені), різко різняться по малюнку борозен; відзначається значна внутрішньовидова (індивідуальна) мінливість; так, наприклад, величина потиличної області кори мозку людини коливалася від 9072 мм до 11748 мм, у орангу - від 4036 до 5350 мм, у нижчих древніх мавп (церкопітек) від 1842 до 2594 мм; 2 - гомологія борозен та звивин у різних тварин зазвичай невідома; 3 - різні автори, незалежно один від одного, дають різні назви тим самим борознам і звивинам; 4 - відсутність єдиної методології дослідження анатомії мозку та єдиної точки зору на морфологічне та функціональне значення борозен. Проте вивчення архітектоніки поверхні півкуль великої кількості видів та груп ссавців дозволило виділити кілька загальних закономірностей анатомічної організації півкуль.
Формування малюнка борозен і звивин відбувається в пре-і постнатальному онтогенезі тварини поступово. Першими формуються головні, первинні борозни і звивини (наприклад, ринальна, гіпокампова, сильвієва), потім вторинні і т. д. З кількістю звивин і борозен тісно пов'язані такі важливі параметри розвитку мозку, як: площа та відносний обсяг, що займає тій або іншою кірковою формацією.
Різні архітектонічні поля зазвичай займають різні звивини. Межі таких зон визначають по дну протилежних звивин, так званих «лімітуючих борозен» (наприклад: аркуатна, центральна борозни у приматів; латеральна, супрасильвієва, хрестоподібна і постхрестоподібна борозни у хижих; цингулярна, ринальна або сильві ссавців).
Багато борозни лежать усередині одного цитоархітектонічного поля, при цьому виділяють так звану «осьову борозну», що займає середину того чи іншого цитоархітектонічного поля. Прикладом таких борозен у приматів є верхня та нижня прецентральна звивина. У багатьох випадках одна й та сама звивина включає ряд цитоархітектонічних полів, межі яких анатомічно визначити неможливо. Прикладом є поля За, 3b, 1, 2 на постцентральній звивині. Різні таламічні, підкіркові, стовбурові, мозочкові, спинальні ядра та центри посилають проекції та отримують реципрокні зв'язки від різних специфічних кортикальних звивин або груп звивин .
Існують значні відмінності в малюнку борозен та звивин на рівні загонів, сімейств, пологів, видів ссавців і навіть на внутрішньовидовому, індивідуальному рівні. Необхідно наголосити, що серед сучасних ссавців є представники, анатомія мозку яких значно відрізняється від форми, характерної для всієї групи. Так, у єхидни (представника нижчих однопрохідних ссавців) півкулі мають незвичайно розвинену систему борозен і звивин, що захоплюють навіть область древньої кори (рис. 112).
Цитоархітектонічна структура нової кори докладно вивчена у багатьох ссавців і має низку загальних рис будови. Неокортекс у більшості зон і полів має типову шестишарову будову. Кожен шар кори характеризується певним набором клітинних елементів, системою аферентних та еферентних зв'язків.
Шар 1(Зональний або молекулярний) розвивається в онтогенезі дуже рано як крайова зона стінки мозку і по суті не належить кірковій пластинці. Основне місце у шарі займає система паралельних волокон, а також нечисленні нейрони різного типу. Основний обсяг шару I займають апікальні дендрити

нейронів нижчих шарів, на які закінчуються аксони корково-кіркових зв'язків, підкіркових нейронів, аксони клітин Кахаля - Ретціуса, клітин Мартіноті. Підраховано, що кожен аксон паралельних волокон утворює до кількох тисяч аксо-дендритних (35%) та аксо-шипикових (66%) дотичних, серійних синапсів.
Шар II(зовнішній зернистий) містить велику кількість дрібних та середніх за розміром зірчастих та пірамідних нейронів. Ширина та клітинна густота шару II у різних полях кори різняться. Основна маса синапсів представлена аксо-дендричними та аксо-соматичними контактами, що розташовуються на тілі нейрона або на проксимальних ділянках дендритів. З'являються складні синаптичні комплекси, самі синапси розподіляються шаром нерівномірно, утворюючи скупчення (т. зв. «синаптичні поля, грона, гнізда»). Кордон з нижчим шаром ІІІ нечітка.
Шар III(пірамідний) складається, в основному, з дрібних та середніх пірамід та зірчастих нейронів різного типу. У шарі також переважають аксо-шипикові контакти (55%). Важливо відзначити, що у шарах II – III з'являється велика кількість (12%) аксо-аксональних контактів, утворених аксонами спеціалізованих гальмівних зірчастих нейронів. Морфологія шару сильно варіює від поля до поля, у зв'язку з чим він часто поділяється на ряд підшарів. У верхній частині шару розташоване аксонне сплетення, утворене розгалуженнями асоціативних аферентів (смужка Кез – Бехтерева).
Шар IV(внутрішній зернистий) складається з зірчастих і піра мідних нейронів і найбільш мінливий у новій корі у ссавців. Саме з будови шару IV нову кору класифікують на гомотипическую кору (поля з добре вираженим, але однорідним структурою шаром) і гетеротипическую кору (де шар сильно варіює). За ступенем варіювання структури шару в гетеротиповій корі виділяють агранулярні поля (шар IV відсутня, наприклад, поля 4, 6 прецентральної області) і гранулярні поля (шар IV поділяється на ряд підшарів, наприклад: поля 17, 18 зорової області). Значні зміни відбуваються і у клітинному складі шару. Переважають аксо-дендритичні контакти (60%), на проксимальних сегментах дендритів, багато аксо-аксональних (12%) та аксо-шипикових (30%) синапсів. Особливість шару IV підкреслюється і тим, що тут закінчується переважна більшість таламічних аферентів, що утворюють ще одне аксонне сплетення в корі - смужку Белларже.
Шари V(Гангліозний) містить популяцію великих і гігантських пірамід і невелику кількість зірчастих нейронів різного виду. У ряді полів цей шар поділяється на ряд підшарів, а його елементи досягають гігантських розмірів (80 – 120 мкм). Це гігантські піраміди Беца у полі 4 прецентральної області або піраміди Мейнерта у полі 17 зорової області.
Шари VI(Поліморфний) складається з дрібних полігональних нейронів і спеціальних клітин (нейрони Мартінноті). Цей шар відрізняє досить складна синаптоархітектоніка: тут зустрічаються синаптичні комплекси конвергентного та дивергентного типів, змішані та реципрокні синапси. Тут переважають аксо-дендритичні синапси.
Внутрішньовидова мінливість у структурі однойменних полів досягає більших величин (до 15%). Міжвидова мінливість, пов'язана зі складними процесами еволюції ссавців та їх ЦНС, значно значніша. Так, дослідження поля 9 (фронтальна область кори) у 16 видів ссавців, включаючи людину, проведену за допомогою методів стереологічного аналізу на ЕОМ, показали, що цитоархітектонічна структура поля значно відрізняється у різних видів по ряду параметрів. Так, товщина кори відрізнялася у вивчених видів більш ніж удвічі (наприклад, у людини порівняно з комахоїдними). Шар I у ряду видів дуже тонкий, особливо у приматів (людини та мавп), тоді як у інших дуже широкий (африканського слона, дельфіна білобочки, морської свині). Дуже сильно варіює щільність нейронів у шарах кори. Такі ссавці, як лемури та мавпи, мають надзвичайно високу щільність нейронів, тоді як у слонів та дельфінів вона дуже низька. Сильно варіюють розміри нейронів у шарах неокортексу. Дрібні клітини одного розміру переважають у корі мозку лемурів та нижчих мавп, середні за розміром нейрони відзначені у комахоїдних та хижих, дуже великі нейрони виявлені в корі мозку хоботних та китоподібних. У вищих приматів і людини варіації розмірів нейронів дуже великі, проте їхньою характерною рисою можна вважати присутність великої фракції дрібних зірчастих нейронів у шарах кори III - IV (звідси термін, що часто застосовується при описі неокортексу приматів, - коніокортськс, пилоподібна кора).
Нейронна структура нової кори ссавців надзвичайно різноманітна та складна. Для неї характерна найбільша серед інших відділів ЦНС різноманітність нейронних типів та різновидів. За однією з класифікацій, розробленою Г. І. Поляковим (Інститут мозку АМН СРСР), у новій корі людини виділяється до 56 різновидів нейронів. Весь нейронний склад неокортексу можна розділити на три великі групи нейронів: пірамідні, непірамідні та перехідні нейрони.

Пірамідні нейронирозташовуються в шарах кори II - VI і характеризуються великою різноманітністю форм та різновидів (рис. 113, а).Для типових пірамід характерно тіло конусовидної форми, від апікального полюса якого відходить апікальний дендрит (або дендрити), що досягає шару I, де дихотомічно розгалужується. Від базальної частини тіла відходить більш менш розвинена система базальних дендритів (від 4 до 16). Основна гілка аксона прямує вниз, виходячи за межі кори та утворюючи систему еферентних проекцій. Частка пірамідних нейронів у різних шарах кори сильно коливається, але загалом становить (залежно від зони та поля кори) від 50 до 90% клітин. Вважали, що з пірамідних клітин характерно сильний розвиток дендритної системи, що відрізняється великим поліморфізмом, тоді як аксонна система менш розвинена і одноманітна. Проте в даний час встановлено, що більшість пірамідних нейронів кори мають поряд зі складною системою дендритних розгалужень і складну систему аксонних розгалужень, що утворюють багату мережу колатералей в межах кори (рис. 113; б, в).Тим самим підтверджується думка на те, що і пірамідні нейрони беруть активну участь в утворенні системи внутрішньомодульних та внутрішньокіркових асоціативних зв'язків.
До групи непірамідних(зірчастих) нейронів входять такі різновиди, як нейрони з висхідним аксоном, клітини з подвійним букетом дендритів з низхідними або висхідними аксонами, клітини з локальними аксонними розгалуженнями, клітини-«канделябри», великі та малі кошикові зірки і шипиків і ряд дрібніших різновидів. До різновидів непірамідних нейронів також відносять ряд спеціалізованих клітин кори: веретеновидні нейрони, клітини Мартінотті, нейрони Кахаля – Ретціуса. Всі різновиди непірамідних клітин складають у корі приблизно 10%, але їхня функціональна роль дуже велика. Саме непірамідні нейрони, їх аксонні терміналі утворюють систему внутрішньокіркових зв'язків, регулюють перебіг фізіологічних процесів у коркових модулях та їх об'єднаннях.
Для великої кількості зірчастих нейронів доведено гальмівну функцію: їх аксони закінчуються на тілах, аксонному горбку, початковому сегменті аксона - місцях, де генерується нервовий імпульс. У синапсах виявлені популяції синаптичних бульбашок овальної форми, що містять гальмівні медіатори: ГАМК(гама-аміномасляна кислота), гліцин, глутамат та ін. б завершують формування структурно-функціональної цілісності кори. Наявність у корі певного набору зірчастих нейронів є важливим показником рівня еволюційного розвитку кори. У нейроморфології навіть використовують спеціальний термін «зірчастість», визначальний рівень структурно-функціонального розвитку кори.
Перехідніформи кіркових нейронів поєднують у собі ознаки того чи іншого основного класу нейронів: наприклад, пірамідо-зірки, зіркоподібні піраміди, пірамідо-веретена і т.п. див. розділ 1).
Детальний якісний та кількісний аналіз структури неокортексу у представників різних груп ссавців дозволив зробити висновок про відносну уніформність, консервативність кори. Так, кількість нейронів в одиниці об'єму кори у різних видів ссавців і в різних зонах кори виявилася однаковою, відношення кількості пірамідних і непірамідних клітин відносно постійно у мавп, кішок і щурів, більшість типів непірамідних клітин, включаючи спеціалізовані види клітин - «канделябрів», зустрічаються у всіх ссавців від комахоїдних до приматів. Отримано дані про подібні гістохімічні та медіаторні параметри неокортексу різних ссавців.
У корі існує певна ієрархія зон, які беруть участь у сприйнятті, аналізі та переробці афферентної інформації різного виду, що надходить в ці зони. В еволюції ссавців у корі відбувається поступове вичленування із загальної, недиференційованої кори специфічних зон. Це так звані проекційні (сенсорні) зони та асоціативні зони. Поля проекційних зон беруть участь у безпосередній переробці стимулів певної модальності (соматосенсорної, зорової, слухової тощо) і поділяються на центральні (первинні, за старою термінологією) та периферичні (вторинні) поля. Наприклад: у людини в соматосенсорній ділянці виділяють центральне поле 3, у зоровій - полі 17, у слуховій - полі 41 і відповідно периферичні поля 1 та 2; 18 та 19; 42 та 22.
Показано, що центральні поля мають більше значення у безпосередньому аналізі первинної сенсорної інформації, а периферичні поля - в інтеграції сигналів тієї ж модальності, які направляються потім у відповідне центральне поле. Центральні та периферичні поля пов'язані одне одним системою двосторонніх зв'язків. Ссавці з маленьким мозком і невеликим неокортекс мають, як правило, лише кілька первинних і вторинних сенсорних і моторних зон, що займають при цьому більшу частину нової кори. Ці райони включають первинні та вторинні соматосенсорні поля SI та S2, первинні та вторинні зорові поля VI та V2, первинну слухову кору AI та первинні моторні поля MI (рис. 114). Ссавці з вищих загонів, що мають більші півкулі і велику площу неокортексу, мають поряд з базовими, основними сенсорними і моторними зонами, велику кількість додаткових сенсорних полів. Так, наприклад, у кішок є щонайменше 7 слухових полів, 12 зорових, 5 соматосенсорних. У мавпи виявлено 6 слухових, 15 зорових та 7 соматосенсорних полів.
Порівнюючи структуру мозку таких ссавців із структурою мозку однопрохідних, сумчастих та нижчих плацентарних, неокортекс яких включає ті ж зони, можна припустити, що подібна базова

hie. 114
Підрозділ зон неокортексу на первинні та вторинні поля у різних ссавців (Kaas. 1992)
а -еЛс. б -білка, в -кішка, г- Мавпа. MI- первинні моторні, SI, SII - первинні та вторинні соматосенсорні, AI- первинні слухові, VI, VII- первинні та вторинні зорові поля. Пояснення у тексті
схема будови неокортексу склалася у примітивних мезозойських ссавців більш ніж 200 млн років тому. Нові зони в неокортекс могли з'являтися з різних джерел: або шляхом вичленування всередині вже сформованих високодиференційованих сенсорних зон, або поступово розвиваючись на основі первинно існуючих недиференційованих зон. У зв'язку з цим вважають, що збільшення різноманітності сенсорних і моторних зон і полів неокортексу могло відбуватися в різних лініях ссавців незалежно і є результатом паралельної або конвергентної еволюції та однойменні поля та зони неокортексу, можливо, негомологічні.
Важливим етапом розвитку неокортексу було вичленування в корі асоціативних (третинних та четвертинних) зон і полів, у яких відсутнє пряме представництво тієї чи іншої чутливості (модальності), а відбувається широка конвергенція різноманітної інформації, що надходить сюди як з первинних сенсорних та проекційних полів з інших зон неокортексу. Асоціативні області кори відрізняються, крім того, найбільш складним та диференційованим клітинним складом, особливо різноманітні у цих полях зірчасті нейрони. У вищих приматів та людини основна маса асоціативних полів зосереджена у лобовій та тім'яно-скроневій областях півкуль.
Асоціативні зони нсокортекс сильно диференційовані і включають кілька груп нулів, кожна з яких має свої характеристики і займає певну ділянку кори. Група I (парасенсорні поля) тісно пов'язана з відповідними первинними та вторинними полями даної модальності, їхня активність залежить від діяльності цих зон. Група II схильна до впливу з боку первинних зон лише частково, оскільки їхня відповідь на аферентні стимули різної модальності формується відносно незалежно від активності первинних проекційних зон. Група III абсолютно незалежна від активності первинних зон, тут відбувається найскладніший синтез різномодальних сигналів, які у цю зону із джерел. Асоціативні зони і поля формуються на останніх етапах пренатального розвитку, які остаточне дозрівання відбувається після народження.
У процесі еволюції ссавців відбувалося послідовне збільшення площі, займаної асоціативними полями та посилення варіабельності у розташуванні однойменних зон кори навіть у близьких родинних тварин (рис. 115). В основному за рахунок збільшення числа та розмірів асоціативних зон відбувається і загальне збільшення площі неокортексу.
Кожне поле та область кори мають численні зв'язки з іншими відділами кори та частинами мозку. Приблизно половина з них утворена аферентними волокнами, що входять у поле. Вони поділяються на дві великі групи: кортико-кортикальні (асоціативні та міжлолушарні) та зв'язки, утворені аферентними волокнами з відділів мозку, що знаходяться нижче. Асоціативні

Рос. 115
Розташування та ступінь розвитку кіркових полів різних типів на латеральній поверхні півкуль ссавців (Богословська, Поляков, 1981) а- їжак, б- щур, в -собака, г- мавпа, д -оранг, е- людина; / - 3 - поля неокортексу: первинні (/), вторинні (2) та третинні (.3); 4 - територія стародавньої, старої та проміжної кори; 5 - нюхова цибулина; 6 - ринальна борозна зв'язку об'єднують поля в межах однієї півкулі. У освіті цих зв'язків беруть участь нейрони багатьох верств кори, у своїй аксони пірамідних нейронів становлять від 60 до 90% зв'язків. Розподіл терміналей асоціативних волокон залежить від шару кори, в якому проходять кортико-кортикальні аксони, а також від типу полів, пов'язаних один з одним. Так, поля, що не мають зв'язків з боку іншого поля із сусідньої півкулі, не утворюють асоціативних зв'язків з однойменним полем і в межах своєї півкулі. Переважна більшість асоціативних волокон закінчується на дистальних ділянках дендритів та його шипиків як пірамідних, і непірамідних нейронів. Є й певні видові відмінності у створенні однойменних асоціативних зв'язків: так, у мавпи поле 7 тім'яної області пов'язано асоціативними зв'язками з меншою кількістю полів, ніж у кішки.
Комісуральні волокна зосереджені переважно у мозолистому тілі, corpus callosum. Його формування пов'язане з розвитком нової кори, проте при різкому збільшенні у вищих ссавців, порівняно з нижчими, площі мозолистого тіла щільність волокон зберігається на одному рівні: приблизно 400 тисяч волокон на 1 мм2 перетину. У корі є ділянки, як утворюють комісуральні зв'язки (їх більшість), і позбавлені їх (поле 17 зорової кори). Більшість калозальних зв'язків є симетричними, хоча є і несиметричні проекції між полями різних функціональних зон кори. Нейрони, що утворюють калозальні зв'язки, розташовуються у всіх шарах кори, з переважною концентрацією в III та V шарах, а їх закінчення виявлено у всіх шарах кори (рис. 116). Виявлено, що ряд нейронів як пірамідного, так і непірамідного типів утворюють одночасно калозальні та асоціативні зв'язки. Морфологічно це забезпечується присутністю таких нейронів розвиненої системи довгих аксонних колатералей. Серед інших трактів, що утворюють міжпівкульні асоціативні зв'язки, необхідно згадати передню комісуру (commissure anterior), добре виражену у всіх ссавців і сполучну структури древньої кори (палеокортексу), а також гіпокампову комісуру (commissure hippocampi), з'єднуючи кори.
Аферентні зв'язки включають, крім перелічених вище, таламо-кортикальні та екстраталамічні проекції. Основна маса специфічних таламічних аферентів проходить у IV шар, хоча і в інших шарах кори також є їх закінчення. На пірамідних

Hue. 116
Організація деяких провідних систем кінцевого мозку ссавців (Йонтов п ін., 1990, Szentagotai, 1983):
а-г-розвиток мозолистого тіла (СС)у їжака («). щури (б), кішки ( в), мавпи (?), О- місця утворення та закінчення калозальних волокон та кори, е-загальна схема організації асоціативних та міжолушарних зв'язків у неокортексі, 1-VI- шари кори, зірочка - гомологічні модулі в сусідніх півкулях, I- Корковий модуль; 2 міжмодульні зв'язки; 3 - зв'язки між зонами в межах півкулі (аксони пірамідних нейронів): 4. Р» -міжпівкульні зв'язки між гомологічними () та негомологічними (5) зонами кори та модулями нейронів кори таламічні аференти утворюють переважно аксо-шипикові синапси, на непірамідних нейронах різного типу - аксо-дендритні та аксо-соматичні. Діаметр зони розгалуження таламічних аферентів становить від 300 до 500 мкм і багато в чому визначає розмір кіркових модулів.
Екстраталамічні аференти беруть початок в основному від центрів стовбура і відповідно до джерела проекції поділяються на холін-, адрен-, дофамін- та серотонінергічні. Ці волокна проходять у всі шари кори (в кожній системі є свої особливості розподілу закінчень по шарах), йдуть паралельно поверхні кори на великій відстані (до декількох міліметрів) і утворюють типові синаптичні з'єднання, так і модуляторні контакти. Загальна кількість таких закінчень становить від 5 до 12% від числа міжнейронних контактів у корі.
Найбільш великими еферентними шляхами вважаються кортико-спінальні (пірамідний тракт) та кортико-бульбарні тракти. Вони утворені аксонами пірамідних нейронів нижніх (V – VI) шарів кори. Більшість еферентів (особливо що йдуть від моторних і сенсорних полів кори) мають суворо топічну організацію. Волокна низхідних трактів формують численні колатералі, що закінчуються у різних відділах мозку. Так, колатералі волокон пірамідного тракту закінчуються на нейронах червоного ядра, що утворюють рубро-спінальні проекції, і впливають на їхню діяльність.
Напрями еволюційного розвитку структури нової кори у різних видів плацентарних ссавців виявилися різними. У ссавців зустрічається кілька варіантів будови неокортексу.
Перший тип неокортексузустрічається у представників таких загонів, як: комахоїдні, рукокрилі, гризуни, зайцеподібні. Неокортекс цих тварин займає від 30 до 60% поверхні півкулі, асоціативні області розвинені дуже слабко. Практично відсутні борозни та звивини (за винятком основних, характерних для всіх ссавців). Для цитоархітектонічної будови кори характерно "переважний розвиток I шару і нижніх шарів V - VI, практична відсутність IV шару (рис. 117). Рівень нейронної диференціювання досить низький. Пірамідні нейрони шару II мають систему віялоподібних дендритів. Система розгалуження дендритів є атиповою.Серед пірамід нижчележачих шарів відзначається велика варіабельність за розмірами і формою дендритного дерева: від типових пірамід дот.н. - III, множини перехідних, недиференційованих форм.Зірчастих нейронів мало, хоча в зародковій формі в корі різних представників цієї групи ссавців можуть зустрічатися всі основні різновиди непірамідних клітин. синапсів асиметричного типу (тип I за Греєм). Синапси з сплощеними синаптичними бульбашками, котрим, часом, доведено гальмівна природа контакту, зустрічаються дуже рідко. Також рідкісні та аксо-аксональні контакти.

Рос. 117
Цитоархітектоніка (а) та нейронна організація ( б) центральної області неокортексу довгокрилої кажанів Minioptcrux sthreibeni
- (Ferrer, 1987)
- 1 -4 - пірамідні нейрони різних верств; 5 - піраміда II шару з широкою зоною розгалуження дендритів; 6-#- гігантські (6 7) і крейда до не (#) мульти полярні радіальні нейрони; 9 біполярний нейрон, /- VI- шари кори

Рис. 118
Зовнішній вигляд півкуль кінцевого мозку дельфіна (Tursiops truncatus)
(Jacobs е. а., 1984)
SSylEct- ектосівнева борозна, орбітофронтальна ( OFOper), тім'яна ( Рорег), скронева (Toper)оперкуляризації (випинання) поверхні півкулі
Міжвидові варіації у площі неокортексу можуть досягати більших величин. Так, у сімействі тенрекові (напівводні комахоїдні) у їжакового тенрека неокортекс займає 18,5% площі всієї кори, а у видрової землерийки - 36,7%; у цьому. Кроти у європейського їжака - 30,9%, у піренейської хохулі - 41,4%. Товщина кори коливається від 0,5 мм у деяких видів кажанів, до 1,8 мм у комахоїдних. При цьому варіації товщини кори в межах одного загону або сімейства також дуже великі (наприклад: у комахоїдних – від 0,4 мм у білозубки малюка) Suncus etruscus,до 1,8 мм у видрової землерийки Patomogale velox).
На структуру мозку нижчих плацентарних впливають адаптивні процеси, пов'язані з розвитком ряду сенсорних систем тварин. Так, у європейського їжака (Erinaceus europaeus)виявлено полісенсорну область кори, яка має складний, диференційований нейронний склад і можна порівняти з асоціативними областями кори вищих ссавців. Як другий приклад можна навести дані по структурі кори мозку білки (зорова зона). У корі сильно розвинені шари П - III, добре виражений шар IV, який поділяється на ряд підшарів, виявлено велику кількість високодиференційованих зірчастих нейронів різного типу. Загалом рівень розвитку зорової кори білки можна порівняти з таким у приматів. Ще одним прикладом такої адаптивної спеціалізації може стати феномен сильного розвитку слухових зон неокортексу у ехолокуючих тварин (кажани).
Другий тип неокортексупредставлений у китоподібних. Своєрідність даного загону ссавців у тому, що вони освоїли водне середовище вдруге, виникнувши від наземних форм древніх ссавців, які входять у одну чи кілька гілок древніх копитних.
Кінцевий мозок китоподібних має дуже своєрідний вигляд (рис. 118). За загальним малюнком борозен і звивин він частково нагадує мозок копитних і хижих, але значно відрізняється сильним розвитком дрібних борозен і звивин, різким вигином (курватурою) тім'яної частини півкуль, практичною відсутністю скроневої частки. Індекс енцефалізації дуже високий (96), а неокортекс займає до 97,8% усієї площі кори півкуль (більше, ніж у вищих приматів та людини!). Крім того, у китоподібних, зокрема у дельфінів, виявився зовсім інший малюнок розподілу первинних моторних та сенсорних зон у півкулях (рис. 119).
Аналіз мікроструктури кори, проте, показав, що загальний рівень організації такого неокортекс досить низький. Маючи велику площу неокортексу, китоподібні за рахунок дуже невеликої товщини кори (1,5 - 2,2 мм) мають дуже низькі об'ємні показники (нижче, ніж у комахоїдних і гризунів). Для цитоархітектонічної будови характерні: велика ширина шарів I і II, сильна диференціація шару III (у зоровій корі дельфінів Stenella coeruleoalbaі Tursiops truncatusу його складі виділено три підшари), практично повна відсутність шару IV, а також сильний розвиток філогенетично давніх верств V – VI. Відзначається слабка варіабельність цитоархітектонічної будови різних зон неокортексу, що надає корі мозку китоподібного «монотонного» вигляду.
Для нейронного складу неокортексу також характерний ряд специфічних характеристик будови (рис. 120). Таламічні субкортикальні аференти, входячи в кору, розгалужуються дихотомічно і закінчуються практично у всіх шарах кори, утворюючи численні аксо-соматичні та аксо-дендритні синапси. Найбільша щільність синапсів спостерігається у верхніх шарах кори, причому більша їх частина відноситься до простих, недиференційованих
Мал 119
Схема розташування основних моторних та сенсорних проекційних зон у неокортексі гіпотетичного предка ссавців та локалізація цих зон у сучасних ссавців (Morgan е.а., 1990):
а -гіпотетичний предок ссавців, б - д -сучасні ссавці: кролик (0). кішка (в), мавпа (/) та дельфін (d). 1 -4 - сенсорні зони: слухова (/), зорова ( 2 ). соматосенсорна (3). моторна (4)


Рис 120
Схема нейронної будови та синаптичних взаємин у зоровій корі неокортексу дельфіна (Morgan е.а., 1990)
I - 6 - різні види пірамідних нейронів, 7.8 - середні (7) та дрібні (8) зірчасті яййрони, а-до- Різні види синапсів, що зустрічаються в неокортексі дельфіна, aff- таламічні аференти, I-V- шари кори
форм; зокрема, дуже багато контактів типу «еп passage*». Шари II і III містять велику кількість пірамідних нейронів нетипової, перехідної форми: перевернені піраміди, піраміди з віялоподібними апікальними дендритами, піраміди-веретена і т. д. Вони посилають свої аксони в білу речовину, але по ходу утворюють численні повернення. дендрит нейронів II III шарів. Подібні пірамідні нейрони зустрічаються в неокортекс нижчих ссавців. У чисельному відношенні пірамідні нейрони переважають. Крім пірамідних нейронів, у неокортексі китоподібних виявлено велику кількість великих радіальних зірчастих нейронів ізодендритного типу. Їх довгі, слаборозгалужені дендрити охоплюють велику площу, а аксонні розгалуження закінчуються на тілах і дендритах нейронів декількох шарів. Клітини такого типу відносяться до різновидів ретикулоподібних нейронів, які є консервативно давніми формами кіркових нейронів, характерними, як вважають, для початкових етапів еволюції нсокортексу ссавців. Високодиференційованих зірчастих нейронів різного типу в неокортекзі китоподібних практично немає, у зв'язку з чим у них немає такого важливого показника, як грануляція кори (коніокортекс).
У цілому нині неокортекс китоподібних поєднує у собі ряд примітивних рис будови, які дозволяють, з теорії еволюції неокортексаФ. Занидеса, розглядати їх неокортекс як структуру, що зупинилася у своєму розвитку на третій (параінсулярно-паралімбічній) стадії. При цьому в мозку дельфінів виявлено систему довгих асоціативних внутрішньокіркових зв'язків, що характерно для кори приматів та людини; їм властивий високий рівень розумової діяльності, складні форми поведінки. Все це свідчить про дуже своєрідний, багато в чому незалежний від інших ссавців, шляхи еволюції кінцевого мозку китоподібних.
Третій тип неокортексусформувався в еволюції таких груп ссавців, як: хижі, ластоногі, копитні, хоботні. Незважаючи на наявні відмінності в організації півкуль у представників цих загонів сучасних ссавців, їх поєднує ряд загальних рис будови неокортексу. Півкулі мають добре розвинену систему великих дугових звивин і борозен, що істотно відрізняють їх від мозку інших ссавців і створюють певні труднощі при зіставленні цитоархітекторних карт мозку (рис. 121). У більшості видів відсутній справжній острівець Рейлі, перипалеокортикальні поля якого розташовуються у хижих практично на поверхні, тоді як у приматів неокортекс, що розростається, відсуває острівець в глиб бічної поверхні півкулі. З цим пов'язана і майже повна відсутність чітко вираженої скроневої частки півкуль. Рельєф кори складніший у великих видів, ніж у дрібних (аж до мікрогірії - утворення численних дрібних третинних і четвертинних борозен і звивин, що різко ускладнюють малюнок кори: наприклад, у ведмедів). Індивідуальна та міжгрупова мінливість у цій групі ссавців дуже велика.
Неокортекс займає від 80 до 95% площі півкулі та має велику товщину (від 2,5 до 3,5 мм). Для нейронної структури, найбільш повно вивченої у представників хижих ссавців (кішок та собак), характерне значне збільшення різноманітності та поліморфізму нейронів. Інтенсивно розвиваються шари III і IV, збільшується кількість зірчастих нейронів, зменшується кількість перехідних форм (зірко-пірамід, пірамідо-веретен і т.д.), відсутні радіальні нейрони та атипові форми пірамід, помітно ускладнюється система

Мал Ш
Рельєф нової кори на латеральній поверхні кінцевого мозку у деяких копитних, мозоленогих та хижих (Богословська, Поляков, 1981)
- - європейський кабян, 6 - нільгау, ~ зубр, г - двогорбий верблюд, д -домашній кінь,
- - ведмідь
дендритних та аксонних розгалужень пірамідних нейронів шарів III та V. У неокортексі намічається поділ на гранулярну (лобова частка) та агранулярну (тім'яно-потиличну та скроневу частки) кору; крім численних первинних та вторинних проекційних та сенсорних полів утворюється складна система асоціативних зон.
Однак було б природним очікувати, що в такій великій групі ссавців є різні варіанти будови нсокортексу. ссавців. Це відносно слабка шаруватість кори, відсутність чітких меж шарів (особливо між III і IV), велика кількість «нетипових пірамід», слабке диференціювання зірчастих нейронів. Таким чином, у рамках даного типу неокортексу ми зустрічаємося з низкою різновидів, що є відображенням складних філогенетичних зв'язків у цих загонах ссавців.
Четвертий тип неокортексусформувався у приматів. Це один із найдавніших загонів ссавців, і не дивно, що саме в межах цього загону рівень міжвидової мінливості дуже великий. Так, коефіцієнт енцефалізації у напівмавп становить 0,13 - 1,3; нижчих вузьконосих мавп – 0,56 – 2,22; широконосих мавп Нового Світу – 0,54 – 8,4; людиноподібних мавп – 2,03 – 7,35; людини – 32. Нова кора займає при цьому від 85 до 96% площі півкулі. У приматів формується особливий малюнок борозен і звивин, основу яких складають великі первинні звивини, що йдуть у поперечному напрямку. Відсутня система дугових звивин і добре розвинена скронева частка (острівець Рейлі знаходиться при цьому в глибині латеральної стінки півкулі). Це значною мірою ускладнює гомологізацію рельєфу борозен, звивин та полів неокортексу приматів та інших ссавців.
У приматів інтенсивно розвиваються асоціативні зони кори, особливо в лобовій та тім'яній області півкуль, максимального розвитку вони досягають у людини. Серед шарів кори у приматів сильний розвиток набувають III та IV шари, найбільш помітно поділ коркових зон на гранулярні та агранулярні поля. Збільшується і різноманітність нервових клітин, особливо зірчастих. При цьому рівень варіабельності в однойменних полях кори у різних представників приматів дуже високий.
Таким чином, дослідження структури нової кори у великої кількості видів плацентарних ссавців практично з усіх основних загонів дозволили зробити низку важливих висновків щодо напрямів її еволюційного розвитку. По-перше, загальний план будови неокортексу єдиний всім ссавців. По-друге, неокортекс приматів (і людини в тому числі) є лише одним із можливих та реалізованих в еволюції типів розвитку неокортексу. По-третє, еволюція нової кори проходила у різних загонах ссавців незалежно, паралельно. І, нарешті, у кожному загоні ссавців зустрічаються представники, рівень розвитку неокортексу яких відповідає типу неокортексу, найбільш характерному для цієї групи.
В даний час створені різні теорії організації неокортексу ссавців, серед яких найбільшого поширення набула модульна концепція структурно-функціональної організації кори.
Нейрони кори організовані в строго впорядковану систему вертикальних об'єднань - модулів різного розміру та форми. Вони вперше були виявлені щодо соматосенсорної кори мозку щурів. Ці об'єднання мають форму вертикальних барило, «barrels*», діаметром від 150 до 400 мкм. У барильці в середньому 4000 нейронів, по периферії щільність клітин вище, ніж у центрі. Основна маса дендритів і аксонів розгалужуються в межах барила. Було показано, що кожній вібрисі на мордочці тварини відповідає певний модуль в корі. Надалі було показано, що більшість зон неокортексу у різних видів ссавців мають модульну будову.
Основною передумовою формування нейронних об'єднань є дендритні пучки, утворені апікальними дендритами груп пірамідних нейронів. Вони були описані в різних областях кори щура, кролика, кішки, мавпи, людини, а також у представників нижчих загонів ссавців: кажанів, їжаків, сумчастих. У дендритних пучках створюються сприятливі умови для дивергенції імпульсів, коли одне еферентне волокно утворює кілька контактів з різними дендритами пучка, так і для конвергенції імпульсів, коли на одному дендриті закінчуються аксони з різних джерел. Отже, дана форма поєднання нейронів у неокортекс є для ссавців універсальною. Кількість дендритів у пучках коливається від 2 – 3 до 20 залежно від виду тварини та зони мозку. Між тілами та дендритами пірамідних нейронів виявлено численні електротонічні контакти, що створюють основу для координованої роботи нейронів.
Число клітин, що входять в елементарний кірковий модуль ссавців, становить приблизно 110 клітин. Кожне таке угруповання формується в онтогенезі з окремих нейрогенетичних локусів у вентрикулярній зоні стінки мозку. Вважають, що у продукуванні нейронів модуля одночасно бере участь не більше 10 клітин. Авторадіографічні дослідження розвитку неокортексу мишей показали, що кожен локус продукує один мітотичний цикл 7-9 нейронів. Враховуючи, що в період розвитку неокортексу миші формується 12 генерацій нейронів, загальна кількість клітин елементарного модуля наближається до 110.
Елементарні модулі поєднуються в макромодулі, кількість клітин у яких становить від 3 до 10 тисяч клітин. Загальна кількість макромодулів у корі ссавців коливається від 0,6 до 2 – 3 млн.
Як зазначалося раніше (див. розділ 1), до складу кіркових модулів входить три групи клітин: «аферентні нейрони, пов'язані із зовнішніми (стосовно модуля) аферентними волокнами різного походження; інтернейрони (до 10 типів), що формують складну систему внутрішньо- та міжмодульних зв'язків; «еферентні нейрони, аксони яких виходять за межі кори. Кожна з цих груп нейронів і навіть окремі його елементи утворюють у складі модуля найскладнішу систему просторово чітко організованих «мікромодулів» різного виду та розмірів.
Розміри коркового модуля багато чому визначаються зоною розгалуження специфічних таламічних аферентів і як від 300 до 500 мкм. Основна маса таламічних аферентів розгалужується на рівні IV шару, де на апікальних дендритах кожного пірамідого утворює по 1 - 2 синапси. Крім пірамідних нейронів, таламічні аференти контактують із зірчастими нейронами різного виду, розташованими в цьому ж шарі (рис. 122).
Модульний принцип організації неокортексу є, очевидно, універсальним всім ссавців (є дані, що з модульному принципу побудовано багато відділів ЦНС як ссавців, а й інших хребетних, як вищих, і нижчих).
Різноманітність модулів у кіркових структурах мозку ссавців досить обмежена. Немає кортикальних зон і полів, які містили б кілька типів модулів. Так, у первинній

Рис 122
Схеманейронної організації «елементарного коркового модуля» (Szentagotai, 1978 - 1984)
1 - пірамідні нейрони; 2 - 6 - різні види зірчастих нейронів; 7.8- специфічні (7) та кортико-кортикальні (8) аференти та зони їх розгалуження, геометричні фігури визначають зони розгалуження відростків клітин модуля та межі самого модуля, I - VI -шари кори зорової кори макаки виявлено всього три типи морфо-функціональних модулів: «колонки окодомінантності», які активуються або правим, або лівим оком; "орієнтаційні колонки", нейрони яких реагують на певне положення предметів у просторі і так звані "цитохромоксидазні колонки", де відсутня перевага нейронів до певної орієнтації стимулу, але віддається перевагу кольору. У складі останніх виділяється два підтипи модулів, пов'язаних із відчуттям жовто-блакитної гами кольорів або червоно-зеленої. У первинній соматосенсорній корі макаки (поле ЗЬ) і кішок виділяється не більше двох типів модулів: один пов'язаний із швидкою реакцією на шкірну стимуляцію, інший – з повільною. Є зони, де виявлено лише один тип модулів (наприклад, у соматосенсорній корі (SI) мозку щурів). Можливо, розвиток методичної бази досліджень дозволить у ряді випадків довести наявність більшого розмаїття модулів у тій чи іншій зоні кори, проте значного збільшення їх розмаїтості, мабуть, не відбудеться. Причини такого еволюційного обмеження не зрозумілі.
Цікаву модель становлення модульної організації в еволюції ссавців запропонував П. Морган із співробітниками. Відповідно до цієї моделі в еволюції відбувалося утворення в корі модулів з їх подальшою модифікацією (ускладненням) та збільшенням кількості. Вихідний (гіпотетичний) тип мозку предків ссавців характеризувався наявністю в корі невеликої кількості легко організованих модулів. У цих модулях

Рис 123
Схема можливої еволюції модульної будови кори ссавців
- (Glezere. а., 1988)
- б -
а- Вихідний гіпотетичний модуль, б- Модифікований модуль. I- піраміди шару II з широкою зоною розгалуження дендритів; 2 - піраміди шарів III – V; 3 - муль-типолярна клітина; 4 - біполярна клітина; 5 – короткоаксонні зірчасті нейрони різних типів; 6 - специфічні та 7- неспецифічні таламо-кортикальні аференти специфічні та неспецифічні таламо-кортикальні волокна, а також асоціативні та комісуральні проекції направлялися переважно до шару I, контактуючи з апікальними дендритами пірамідних і (3) непірамид. Великі мультиполярні ізодендритні нейрони посилали свої аксони на другий шар, забезпечуючи внутрішньокіркові зв'язки. Таким чином, основним «аферентним» шаром кори був шар II з його спеціалізованими пірамідними клітинами, аксони яких прямували в шари кори, що все нижчележать.
У процесі еволюції у модулях відбувався поділ зон закінчень основних аферентних проекцій. Таламо-кортикальні аференти закінчуються переважно в новому шарі IV, що інтенсивно розвивається, де з'являється велика кількість високоспеціалізованих зірчастих нейронів. Інші аференти прямують у верхні шари кори, даючи на протязі численні колатералі.
У різних групах ссавців процес модифікації та збільшення кількості модулів йшов, на думку автора цієї гіпотези, незалежно, внаслідок чого до теперішнього часу сформувалося чотири основні типи неокортексу. Консервативний, при якому зберігаються багато ознак гіпотетичного типу кори, зустрічається у представників сучасних загонів комахоїдних та рукокрилих. Прогресивно-консервативнийтип, що характеризується деяким збільшенням кількості модулів та незначним їх ускладненням. Тварини з таким типом неокортексу зустрічаються практично у всіх загонах сучасних ссавців (наприклад, лемури та мавпи серед приматів). При прогресивномуТип будови кори кількість, ступінь складності і різноманітність модулів значно збільшується. У кожному загоні ссавців є представники, неокортекс яких має риси прогресивного типу (серед приматів - це мавпи і людина). Консервативно-прогресивнийтип відрізняється різким збільшенням кількості модулів у корі при незначній їхній модифікації, що призводить до формування «монотонної» структури кори (китоподібні).
Старі кори (archicortex). Стара кора мозку ссавців утворена в основному структурами гіпокампу, який поділяється у плацентарних ссавців на три відділи: прекомісуральний (прекалозальний), супракомісуральний (супракалозальний) та ретрокомісуральцій (посткалозальний). Через сильний розвиток у вищих ссавців мозолистого тіла перші два відділи у них редукуються (рис. 124). Прекомісуральний відділ, розташований між зоною переднього нюхового ядра та мозолистим тілом, представлений дрібними пірамідними клітинами та пов'язаний аферентними та еферентними проекціями с.піриформною та ентерінальною корою. Середня частина гіпокампу перетворюється на вузьку смужку сірої речовини, що зв'язує передній та задній відділи гіпокампу. Ретрокомісуральний відділ найбільш розвинений та представлений структурами субикулуму (S) та власне гіпокампа, що складається з ряду зон (СА 1 - 4) та зубчастої фасції (FD). Гіпокамп має кіркову структуру і складається з п'яти шарів, основним з яких є шар III, утворений пірамідними нейронами. У нижчих ссавців цей шар вузький, компактний; у вищих – широкий, дифузний.
Гіпокамп - найважливіший компонент лімбічної системи кінцевого мозку, пов'язаний з багатьма відділами головного мозку. Він отримує аферентні проекції від різних зон неокортексу (включаючи асоціативні поля), а також септуму, амігдали, ряду ядер таламуса, багатьох відділів стовбура мозку. Еферентні проекції гіпокампу утворені аксонами виключно пірамідних нейронів і проходять переважно у складі склепіння. Через нього забезпечується зв'язок гіпокампа з амігдалою, таламусом, гіпоталаму-

РШС 124
Організація гіпокампу у їжака (а) та людини (б)і будова гіпокампу лемура на тангенціальному зрізі (я)
(Stephan, Malonescu, 1980)
/ - прекалозальний; 2 - супракалозальний та 3 - посткалозальні відділи гіпокампу; 4 - мозолисте тіло; 5 – передня комісура; СА1 і СА2/3 - відділи гіпокампу, ERC - енторинальна кора. FD – зубчаста фасція. PaS – парасубікулум, PrS – пресубікулум, S – субикулум сом, палеокортексом, септумом. У вищих ссавців над мозолистим тілом формується пучок волокон – пояс, cingulum, через який гіпокамп пов'язаний з іншими відділами архікортексу та рядом полів неокортексу. Комісуральні зв'язки здійснюються через комісуру гіпокампу.
Стародавня кора (paleocortex). Стародавня кора у ссавців займає в півкулі вентральне положення, відтіснена з латеральної поверхні новою корою, що сильно розрослася. Для неї характерні слабка шаруватість та неповне відокремлення від сусідніх клітинних мас стріатуму. Основна частина палеокортексу - піріформна кора - отримує нюхові проекції по латеральному нюховому тракті, волокна якого закінчуються на апікальних дендритах пірамідних нейронів піриформної кори. Аксони цих нейронів прямують у нюховий горбок, а також у префронтальну ділянку кори. Палеокортекс має еферентні: зв'язки з амігдалою та епіталамусом. Привертає увагу той Ьакт, що у тварин, що мають редуковану нюхову систему (наприклад, дельфіни, деякі птахи), палеокортекс добре добре. Це свідчить про те, що палеокортекс функціонально пов'язаний не тільки з нюхом.
Супаліальні формації. У кінцевому мозку ссавців: упаліальні формації включають: хвостате ядро, nucl. audatus, і шкаралупу, putamen, що часто розглядаються як єдина: структура (неостріатум), а також бліда куля, globus pallidus, (пагеостріатум), огорожу, claustrum, амігдалу, структури вентромеді- 1ЛЬної стінки півкулі ди- тональної зв'язки Брока, безіменна субстанція, nucl. accumbens).
Цитоархітектонічна та нейронна організація стріатуму 1лекопитающих (він включає формації нео-, палеостріатуму та граду) дуже складна. Неостріатум цитоархітектонічно неоднорідний. Його клітини утворюють складні просторові комплекси - стріосоми, оточені більш рихло розташованими нейронами, що становлять так званий матрикс. На поперечних різах мозку до складу центральної частини стріосоми входить від 1500 до 15000 нейронів різного розміру та форми. Нейрони часто розпагаюються дуже щільно, що передбачає наявність електротонічних онтактів та синхронізації роботи груп нейронів у стріосомах. Форса цих острівців, в основному округла або еліпсоїдна (300 - 00 мкм у діаметрі), проте об'ємна реконструкція стриосом на саітальних і тангентальних зрізах мозку показала дуже складну конфігурацію рослин подібних комплексів (рис. 125).

Мал. 125
Тримісна реконструкція просторових комплексів (стріосом) у хвостатому ядрі ссавців (Goldman - Rakic, 1982) а- Зона матриксу, б -зона стріосом
Матрикс представлений скупченнями рихло розташованих нейронів. Відзначені й суттєві відмінності у медіаторній мозаїці стріосоми та матриксу. Стріосоми хвостатого ядра можна розглядати як аналоги коркових модулів неокортексу.
У складі хвостатого ядра виділено кілька типів нейронів, що розрізняються за розміром (великі, середні, дрібні), за типом аксонів (довго-і короткоаксонні) і характером дендритних розгалужень (густо-і рідковетвисті; шипикові та безшипикові). Показано, що клітини будь-якого розміру можуть бути як довгоаксонними, що утворюють еферентні проекції, так і короткоаксонними, що формують внутрішньостріаторні зв'язки. Великі довгоаксонні нейрони дають висхідні проекції в кору, тоді як дрібні та середні довгоаксонні нейрони - низхідні проекції. В еволюції ссавців частка дрібних нейронів різного типу збільшується.
Для мікроструктури неостріатуму характерна значна кількість дотичних синапсів. Більшість із них - аксо-денд-ритні. Аксони йдуть не паралельно дендритам, а перпендикулярно, перетинаючи один за одним. Незважаючи на деяке переважання дивергенції закінчень, у значній кількості зустрічаються і синаптичні комплекси конвергентного типу, коли на одному дендриті закінчується кілька аксонних закінчень. Ще одна особливість хвостатого ядра - висока щільність нейропіля, утвореного великою кількістю мієлінізованих та немієлінізованих волокон. Більшість цих сплетень становлять колатералі короткоаксонних інтернейронів, і навіть які входять у ядро аферентних волокон. Аксони більшості нейронів закінчується в межах неостріатуму і лише незначна кількість нейронів утворюють зовнішні зв'язки. У зв'язку з цим однією з особливостей будови неостріатуму є велика кількість внутрішніх зв'язків.
Блідий шар поділяється цитоархітектонічно на внутрішній і зовнішній сегменти. У приматів і людини зовнішній і внутрішній сегменти блідої кулі тісно прилягають один до одного, розділяючись тонким шаром волокон, тоді як у більшості інших ссавців гомолог внутрішнього сегмента, відомий під назвою ентопедункулярного ядра (nucleus entopeduncularis), топографічно значно відокремлений .
Нейронний склад блідої кулі досить одноманітний. Його клітини - в основному великі рідкісні нейрони ретикулярного типу зі строго впорядкованою орієнтацією дендритів, що утворюють систему мікромодулів у вигляді дисків, паралельних один одному і латеральній межі ядра, але розташованих перпендикулярно напрямку ходу аксонів нейронів неостріатуму. На противагу хвостатому ядру аксонні розгалуження нейронів блідої кулі йдуть паралельно, утворюючи своєрідну аксо-дендритну систему - тип синапсів, характерних для клітин Пуркіньє мозочка. Часто спостерігаються численні синапси, утворені 6-8 аксонними терміналями. Аксони клітин хвостатого ядра та шкаралупи, проходячи в бліду кулю, пронизують сотні дендритних модулів та утворюють складні синаптичні комплекси.
Стріатум ссавців має складну систему зв'язків із великою кількістю мозкових структур (рис. 126). Основною тенденцією в організації аферентних та еферентних проекцій стріатуму є формування кільцевих зв'язків: неокортекс неостріатум неокортекс, неокортекс неостріатум -? палеостріатум таламус -» неокортекс, неостріатум палеостріатум -> неостріатум,

неостріатум палеостріатум -> таламус -? неостріатум, стріатум -> чорна субстанція -> стріатум. Багато з цих зв'язків топографічно впорядковано. Наприклад, у разі кірково-неостріаторних зв'язків нейрони глибоких шарів кори (V, VI) із певної зони неокортексу проектуються до стріосом, верхніх (II, III) – у матрикс. Є відмінності у переважанні певних типів проекцій від різних зон кори. Еференти від алокортикальних ролей закінчуються в основному в центральній частині стріосоми, тоді як походять з неокортикальних полів - в матриксі. При цьому входи з різних полів кори в неостріатумі можуть перекриватися, що створює основу конвергенції на нейронах стріатуму різноманітної інформації.
Таким чином, філогенетично молода система зв'язків стріатуму з кірковими формаціями отримує у ссавців подальший розвиток, забезпечуючи йому роль одного з інтеграційних центрів кінцевого мозку.
Огорожа - тонка смужка сірої речовини, відокремлена від шкаралупи пучками волокон, що утворюють зовнішню капсулу, capsula externa, а від кори її відокремлює вузька смужка білої речовини - зовнішня капсула, capsula extrema. Топографічно огорожа ділиться на дорсальну, вентральну та каудальну частини. Філогенетично огорожа є новим утворенням базальних гангліїв півкуль; у їжаків та деяких гризунів гомолога огорожі ще немає. Для нейронної будови огорожі характерна значна поліморфність. Найбільш численні кіркові виходи із різних зон неокортексу. Аференти також надходять зі стріатуму, амігдали, таламуса та чорної субстанції. Еференти огорожі прямують до нової кори, стріатуму, амігдалі, нюхової цибулини, таламусу та мозочка. Є дані, що огорожа має кіркове походження.
У септальній зоні кінцевого мозку ссавців, що включає медіальне та латеральне септальні ядра, ядро діагональної зв'язки Шлюбу, nucl. accumbens, і функціонально пов'язаної з лімбічною системою, з'являється, на відміну від аналогічної зони кінцевого мозку рептилій та птахів, велика кількість високоспеціалізованих короткоаксонних нейронів. Проте аналіз зв'язків, структури та гістохімічних характеристик дозволяє проводити гомологію основних відділів септальної зони мозку рептилій, птахів та ссавців.
Нюховий горбок має шарувату будову (розрізняють зовнішній шар волокон, середній шар пірамідоподібних нейронів та внутрішній шар поліморфних клітин). Ступінь вираженості шарів у різних видів ссавців значно варіює. Аферентні проекції надходять із нюхової цибулини, піриформної кори, амігдали, преоптичної області, ряду зон неокортексу, інтраламінарних ядер таламуса, чорної субстанції. Еференти направляються в септальну зону і далі в гіпокамп, стріатум, бліду кулю, дорсомедіальну ділянку таламуса та епіталамус, амігдалу та піриформну кору. Таким чином видно, що нюховий горбок ссавців пов'язаний не тільки з нюхової, але і з лімбічною та стріопалідарною системами.
Амігдала ссавців складається з великої кількості (до 10) ядер, які об'єднуються в декілька груп: базолатеральну (латеральне, основне та додаткове ядра) - найбільш філогенетично молоду частину амігдали, пов'язану з кірковими формаціями; кортикомедіальну (періамігдалярну кору, середнє і заднє кортикальні ядра, медіальне ядро) - філогенетично більш стару частину, пов'язану з нюхової системою; центральну (центральне ядро, передньоамігдалярна зона) - пов'язану з гіпоталамусом та ядрами стовбура мозку, що контролюють вегетативні функції організму.
У філогенезі ссавців в амігдалі відбувалося два процеси. По-перше, зменшувалася частка, яку займали ядри кортикомедіальної групи, і збільшувався обсяг базолатеральної групи ядер (остання досягає найбільшого розвитку та максимальної складності у приматів і людини). По-друге, у вищих ссавців унаслідок сильного розвитку нової кори амігдала зміщується з вентральної стінки півкулі на вентромедіальну. Багато питань організації амігдалярного комплексу кінцевого мозку вищих хребетних ще вирішено. Однак можливо, що амігдалоїдний комплекс, будучи важливою частиною лімбічної системи мозку, бере участь у регуляції багатьох комплексних поведінкових реакцій вищих хребетних.
- В даний час дослідники на підставі гістохімічних, ходологічних та ембріологічних даних відносять перелічені тут структури (починаючи з нюхового ядра) до субпаліальних формацій.
- Є пряма кореляція між розміром мозку та ступенем розвитку звивин - великі півкулі мають, як правило, більш розвинену систему борозен та звивин.
 (1оцінок, у середньому: 5,00із 5)
(1оцінок, у середньому: 5,00із 5)