Заднее ядро трапециевидного тела. Варолиев мост — основная взаимосвязь между отделами мозга. Какие функции он выполняет
Головной мозг человека занимает ключевую позицию в регулировании всех систем организма человека. С помощью этого органа осуществляется связь между деятельностью органов и всех систем. Без мозгового координирования человек не может существовать.
Благодаря изначально настроенной работе ЦНС мы можем передвигаться, говорить и выполнять множество других функций.
Головной мозг человека имеет самое сложное строение, а каждый из его отделов отвечает за свои функции. Таким образом, все структуры мозга поддерживают работу организма в целом.
К основным отделам головного мозга относится непосредственно варолиев мост. Он содержит такие необходимые для жизни человека центры как:
- Сосудистый
- Дыхательный
Также именно он изначально формирует большинство черепных нервов.
Ключевым компонентом главного функционирующего органа представляется нейрон. Она отвечает за прием, обработку и хранение данных. Весь человеческий мозг буквально заполнен данными клетками и их отростками, которые обеспечивают передачу сигналов в органы. Также в состав головного мозга входят серое и белое вещество.
Ключевой структурной частью головного мозга выступают:
- Правое и левое полушарие (Отвечают за нашу память, мыслительные процессы, воображение)
- Мозжечок (координирует и формирует нашу двигательную систему). Именно благодаря мозжечку мы можем передвигаться, чувствовать равновесие, положение тела
- Варолиев мост

Строение варолиева моста
Строение моста с наружной стороны представляется в виде валика, в состав которого входят черепные нервы, артерии, ретикулярная формация и нисходящие пути. С внутренней стороны представляется половинкой ромбовидной ямки.
По срединному пути проходит базилярная борозда, по бокам которых располагаются пирамидные возвышения. Если сделать поперечный срез, то на клеточном уровне можно увидеть белое вещество.
В боковом отделе располагаются ядра верхней оливы, а именно в области переднего основания и задней покрышки. Между данными частями идет линия, которая представляется многочисленными волокнами. Специалисты выделяют данное множественное скопление волокон как трапециевидное тело, которое отвечает за образование слухового пути.
Граница, которая отделяет мост и среднюю ножку мозжечка, называют область, где ответвляется тройничный нерв.

Функции
Мост головного мозга обеспечивает ряд важных функций для человеческого организма, а именно:
- Обеспечивает целенаправленный контроль над движениями тела
- Позволяет воспринимать тело в пространстве
- Контролирует чувствительность языка, кожных покровов лица, слизистой оболочки носа и глазной оболочки
- Отвечает за мимику и слух
- Координирует весь акт потребления пищи (глотание, слюноотделение, пережевывание)
Рефлекторная функция, которую выполняет мост, позволяет человеческой ЦНС отвечает на различные внешние раздражители (рефлекс). Рефлексы подразделяются на 2 вида:
- Условные, которые приобретаются в процессе жизнедеятельности с возможностью корректировки
- Безусловные, которые не поддаются сознания и закладываются в момент рождения (жевательные, глотательные и другие рефлексы)
Также мост выполняет функцию обеспечения взаимосвязи коры головного мозга и подлежащими образованиями. Непосредственно сами волокна направляются к мозжечку, спинномозговому и продолговатому отделу. Данный переход возможен благодаря прохождению через мост нисходящих и восходящих путей.
Все важные функции моста достигаются с помощью черепных нервов.
Например, 5-ая пара черепных нервов отвечает за восприятие боли и тактильных ощущений, а также обеспечивается акт жевания. Отводящие нервы имеют в своем составе двигательные волокна, которые и предоставляют возможность поворота глаз. Также от моста зависит работа дыхательного центра продолговатого отдела мозга.

Патологические состояния
Стоит выделить, что один из ключевых отделов головного мозга, мост, а также мозговые ножки поражаются намного чаще, чем тот же продолговатый мозговой отдел. Зачастую находятся в патологическом состоянии вследствие эмболии, артрита или тромбоза. В данных местах чаще всего возникают кровоизлияния, опухолевые образования, инфекции, например, туберкулы.
Наличие таких патологий достаточно тяжело диагностируется зачастую специалисты устанавливают точный диагноз с помощью дифференцированной диагностики от случая к случаю. Однако на сегодняшний день существуют основные синдромы, которые выделяются определенной клинической картиной.
Головной мозг и мост выделяется следующими типами синдромов:
- Нижний синдром моста
Является самым ранней установленной патологией. Располагается на всей вентральной части перереза Варолиева моста в нижних его отделах. В данном случае наблюдается следующая клиническая картина:
- Гемиплегия центрального вида
- Периферический паралич лицевого и отводящего нервов, также чаще всего поражение парных нервов, располагающихся на противоположной стороне, то есть на стороне очага поражения
- Гемианестезия, когда лицевые нервы поражения на пораженной стороне, а тело и конечности на противоположной
- В редких случаях гемихорея и гемиатаксия
- Верхний синдром моста или синдром Реймона-Сестана
Патология локализуется в задне-боковой части моста, а патологические проявления следующие:
- Незначительный гемипарез без явной изменчивости сухожильных и кожных рефлексов
- Гиперкинезы - атетоз, тремор
- Дизартрия
- Вертикальный нистагм
- Частые головокружения
Т кани головного мозга представлены широким перечнем образований. По своему строению это едва ли не самая сложная часть человеческого организма, что обуславливает широкий характер деятельности центральной нервной системы. При оценке строения можно выделить несколько областей ЦНС в этой локализации.
В основании церебральных структур расположен так называемый ствол головного мозга. Он обеспечивает группу жизненно важных функций: от дыхания и сердечной деятельности и до терморегуляции. Любые повреждения или нарушения работы приводят к тяжелой инвалидности или смерти.
Варолиев мост - это составная часть ствола, расположенная между средним и продолговатым мозгом, которая обеспечивает нормальную проводимость нервных импульсов и дает возможность выполнять ряд произвольных действий.
Отвечает за некоторые функции высшей деятельности. Его повреждения, например, на фоне травмы, инсульта, приводят к критическим нарушениям работы всего организма.
Диагностика поражений этой анатомической структуры представляет определенные сложности из-за глубокой и «неудобной» локализации. Единственным надежным способом обследования считается МРТ или реже компьютерная томография.
Варолиев мост располагается на уровне ствола головного мозга и условно в его анатомии выделяют две основных области.
- Верхняя часть. Состоит из серого вещества и включает в себя несколько пар черепных нервов (с 5-го по 8-ой). Это собственно функциональная структура.
- Нижняя или основание - обеспечивает проводимость сигналов, выступает транспортным путем для естественных импульсов.
На уровне верхней части моста расположена ретикулярная формация. Она представлена большим скоплением волокон, которые позволяют всей ЦНС функционировать слаженным образом.

В основании расположен плотный слой толстых проводящих тяжей. Они представляют собой по три ножки с каждой стороны, соединены с мозжечком и дают возможность работать экстрапирамидной системе.
Ниже Варолиева моста находится продолговатый мозг, в верхней части - средний.

Есть некоторые отличия в структуре названного образования у детей и взрослых. При оценке у пациентов до 8 лет обнаруживается полное отсутствие миелиновой оболочки, что считается нормальным явлением.
Оформление этой части головного мозга наблюдается в ранний подростковый период. Анатомия моста сложная, что объясняется необходимостью осуществлять ряд действий со стороны ЦНС.
Какие функции выполняет мост
Мост головного мозга отвечает за несколько важных форм активности.
Среди таковых:
- Рефлекторные автоматические и произвольные движения глаз и барабанной перепонки в ответ на громкие звуки, также тканей ротовой полости (небо). Любое нарушение заканчивается проблемами.
- Способность к целенаправленной двигательной активности. Поскольку мост в мозге обеспечивает функционирование мозжечка, при любом повреждении наблюдаются проблемы с возможностью управлять телом.
- Восприятие вестибулярных раздражителей. В данном случае речь идет о способности воспринимать свой организм как единой целое, его ориентацию и расположение в пространстве, отвечать на любые изменения окружающих условий, также гасить лишние движения (например, при резком торможении в общественном транспорте, спотыкании и пр.). При поражении отмечается нарушение координации. Возможности ориентироваться в пространстве.
- Обеспечение обонятельной функции. Мост обладает этой способностью частично. Также за нее отвечают другие подкорковые скопления.
- Нормальная иннервация кожи и слизистых оболочек лица.
- Варолиев мост участвует в формировании сна. Это сложная и слаженная работа сразу нескольких церебральных образований. Любые нарушения немедленно приводят к проблемам с ночным отдыхом. Пациент становится вялым, проявляются астенические процессы.
- В функции Варолиева моста входят акты жевания и глотания. Жизненно необходимые для питания, дыхания.
- Собственно от работы этой структуры зависит и способность организма к нормальному газообмену. В отсутствии адекватного проведения импульсов начинаются проблемы, вплоть до расстройств летального характера.
Базовые действия выполняются нервными тканями постоянно. Даже незначительные изменения заметны сразу же.
Варолиев мост является частью ствола головного мозга, потому отклонения в его деятельности становятся косвенной причиной дисфункции всего этого образования.
Вероятны осложнения вплоть до стремительных, смертельных. Качественная медицинская помощь возможна не всегда из-за сложной локализации структур и непростого строения.
Патологии, нарушающие функции моста и их симптомы
Есть группа заболеваний, для которых типично нарушение нормальной работы организма в результате деструкции рассматриваемых тканей.
Среди таковых:
Синдром Бриссо-Сикара
Сопровождается расстройством активности черепных нервов. Определяется односторонним парезом или полным параличом половины тела.
Также утрачивается способность управлять мышцами лицевой области, возможен птоз (опущение века) с нарушением зрительной функции.

Встречается такое расстройство на фоне инфекционного, аутоиммунного или опухолевого поражения. Реже становится итогом . После транзиторной атаки или собственно полноценного инсульта.

Синдром Боннье
Характеризуется поражением группы черепных нервов. В данном случае в конечном итоге страдают слуховые и вестибулярные ядра.
Симптоматика неспецифична. Возникают проблемы с восприятием звуковых раздражителей. Больные постоянно испытывают головокружение, тошноту, слабость.
Составной частью клиники является и бессонница. Пациент становится раздражительным, отмечается нестабильность эмоционального фона. Вплоть до резких смен фаз, как, например, при биполярно-аффективном психозе.
Синдром Грене
Типичная черта этого патологического процесса - нарушение чувствительности мимической мускулатуры, что в конечном итоге приводит к проблемам с проявлением невербальных сигналов, эмоций.
Наблюдается частичный парез жевательных мышц с одной стороны. С другой нарушение управляемости также присутствует, но в значительно меньшей мере.
Вентральный синдром
Крайне сложное состояние. Для него характерна как минимум утрата речевой функции. Это самый легкий случай.
Классическая же ситуация определяется полной потерей способности двигаться. Человек не может пошевелиться. Общение возможно только с помощью глаз.
Такое расстройство сохраняется длительное время. Быстро приводит к застойным явлениям, смерти больного. Восстановление не представляется возможным .
Синдром Раймона-Сестана
Характеризуется ключевым проявлением со стороны глазодвигательных нервов. Человек утрачивает способность произвольно фокусировать взгляд и переводить его с одного объекта на другой.
Возможно спонтанное облегчение состояние и его последующее возвращение по неясным причинам.
Синдром Гюблера
Не имеет специфических проявлений крови паралича мимической мускулатуры. Выражение лица характеризуется как маска.
Пациент не способен адекватно невербально проявлять эмоции и реагировать на окружающие стимулы.
Чувствительность кожи также падает, что обнаруживается по результатам функциональных тестов и физикального исследования.
Синдром Фовилля
Присутствует паралич мимической мускулатуры и косоглазие с расстройством зрения.
Болезнь Гасперини
Сочетанный патологический процесс. Характеризуется смешанной симптоматикой.
Заболевания, приводящие к развитию синдромов
Строение Варолиева моста предполагает множество возможных поражений и столь же большое количество проявлений. Однако есть группа заболеваний, которые становятся фундаментом для названных выше синдромов.
Сюда можно отнести:
- Инсульт. Острое нарушение мозгового кровотока в той или иной области с отмиранием нервных тканей и утратой части функций церебральных структур. Если страдает сам ствол головного мозга, в самом благоприятном случае это закончится нарушением высшей деятельности.

- . Неверно называются микроинсультами. Наблюдается то же самое, но отсутствует значительное отмирание тканей.

- . Нарушение проходимости артерий в результате закупорки таковых холестериновыми бляшками или спонтанного сужения на фоне, например, длительного курения, гипертензии (роста давления).

- Инфекционные процессы. Особенно те, что затрагивают церебральные ткани. Энцефалит, менингит.
- Демиелинизация. Рассеянный склероз.
Варолиев мост отвечает за массу важных функций и имеет системное строение. Лечение патологических состояний, когда деятельность этой структуры уже нарушена, представляет крайне сложный, а порой и невозможный процесс.
Потому имеет смысл превентивно воздействовать на все заболевания, которые могут стать источником проблемы в будущем. Это важная мера профилактики.
Варолиев мост выполняет двигательные, сенсорные, интегративные и проводниковые функции. Важные функции моста связаны с наличием в нем ядер черепных нервов.
V пара – тройничный нерв (смешанный). Двигательное ядро нерва иннервирует жевательные мышцы, мышцы нёбной занавески и мышцы, напрягающие барабанную перепонку. Чувствительное ядро получает афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, 2/3 языка, надкостницы костей черепа, конъюнктивы глазного яблока.
VI пара – отводящий нерв (двигательный), иннервирует прямую наружную мышцу, отводящую глазное яблоко кнаружи.
VII пара – лицевой нерв (смешанный), иннервирует мимические мышцы лица, подъязычную и подчелюстную слюнные железы, передает информацию от вкусовых рецепторов передней части языка.
VIII пара – преддверно-улитковый (чувствительный). Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная его часть – в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
В мосте располагается пневмотаксический центр, запускающий центр выдоха продолговатого мозга, а также группа нейронов, активирующих центр вдоха.
Через мост проходят все восходящие и нисходящие пути, связывающие мост с мозжечком, спинным мозгом, корой больших полушарий и другими структурами центральной нервной системы. По мостомозжечковым проводящим путям через мост осуществляется контролирующее влияние коры полушарий большого мозга на мозжечок.
2. 3. 3 Мозжечок (cerebellum). Мозжечок (малый мозг) располагается сзади от моста и продолговатого мозга. Он состоит из средней, непарной, филогенетически старой части – червя – и парных полушарий, свойственных лишь млекопитающим. Полушария мозжечка развиваются параллельно с корой больших полушарий и достигают у человека значительных размеров. Червь с нижней стороны погружен глубоко между полушариями; верхняя его поверхность переходит в полушария постепенно (рис. 20).
В целом мозжечок имеет обширные эфферентные связи со всеми двигательными системами стволовой части мозга: кортикоспинальной, руброспинальной, ретикулоспинальной и вестибулоспинальной. Не менее разнообразными являются и афферентные входы мозжечка.
Со стволовой частью мозга мозжечок соединяется тремя парами ножек. Самые толстые средние ножки расширяясь, переходят в варолиев мост. Верхние ножки начинаются в зубчатых ядрах мозжечка (см. ниже) и направляются к четверохолмию среднего мозга. Третья пара ножек (нижняя) спускается вниз, сливаясь с продолговатым мозгом. Афферентные волокна, приходящие в мозжечок, преимущественно входят в состав средних и нижних ножек, тогда как эфферентные собраны главным образом в верхних ножках мозжечка.
 Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины; группы извилин формируют дольки мозжечка. Каждую дольку обозначают как классическим названием (язычок, центральная, вершина и т. д.), так и латинской нумерацией (I – Х) в соответствии с распространенной номенклатурой.
Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины; группы извилин формируют дольки мозжечка. Каждую дольку обозначают как классическим названием (язычок, центральная, вершина и т. д.), так и латинской нумерацией (I – Х) в соответствии с распространенной номенклатурой.
Полушария и червь мозжечка состоят из лежащего на периферии серого вещества – коры – и расположенного глубже белого вещества, в котором заложены скопления нервных клеток, образующие ядра мозжечка – ядра шатра, шаровидные, пробковидные и зубчатые (рис. 21).
 Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Она представлена тремя слоями (рис. 22). Самый поверхностный слой – молекулярный
, он состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзинчатых клеток, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье. Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Она представлена тремя слоями (рис. 22). Самый поверхностный слой – молекулярный
, он состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзинчатых клеток, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье. Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Во втором слое коры – ганглиозном – сосредоточены тела клеток Пуркинье. Эти крупные клетки ориентированы вертикально по отношению к поверхности коры мозжечка. Их дендриты поднимаются вверх и широко ветвятся в молекулярном слое. Дендриты клеток Пуркинье содержат множество шипиков, на которых образуют синапсы параллельные волокна молекулярного слоя. Аксоны клеток Пуркинье спускаются к ядрам мозжечка. Часть из них заканчивается на вестибулярных ядрах. Аксоны клеток Пуркинье представляют собой практически единственный выход из коры мозжечка.
Под ганглиозным слоем лежит гранулярный (зернистый) слой, который содержит большое число тел клеток-зерен, или гранулярных клеток. По некоторым подсчетам их число может достигать 10 млрд. Аксоны клеток-зерен поднимаются вертикально вверх в молекулярный слой и там Т-образно ветвятся. Ветви идут параллельно поверхности коры и образуют синапсы на дендритах других клеток. Здесь же в гранулярном слое лежат клетки Гольджи, аксоны которых подходят к клеткам-зернам.
Все клетки коры мозжечка являются тормозящими за исключением клеток-зерен, которые оказывают возбуждающее воздействие.
В мозжечок поступает два типа волокон. Это, во-первых, лазающие, или лиановидные , идущие из нижних олив продолговатого мозга. Нижняя олива получает афферентные импульсы от кожных рецепторов, мышц, суставных оболочек, надкостницы по так называемым спинномозжечковым трактам: заднему (дорсальному) и переднему (вентральному). Лазающие волокна широко ветвятся и подобно лианам оплетают дендриты клеток Пуркинье, формируя на них синапсы. Возбуждая клетки Пуркинье, они усиливают тормозное влияние этих клеток на ядра моста.
Вторая система афферентных волокон – это мшистые, или моховидные волокна, идущие от ядер моста (в которые также поступает информация от рецепторов мышц, сухожилий, вестибулярных ядер) и оканчивающиеся на клетках-зернах. Клетки-зерна посылают нервные импульсы на корзинчатые, звездчатые клетки, клетки Гольжи, возбуждая их, что приводит к торможению активности клеток Пуркинье (снятию тормозного влияния коры на ядра мозжечка).
Таким образом, путь, идущий на клетки Пуркинье через лиановидные волокна, пособствует усилению тормозного влияния клеток Пуркинье на ядра мозжечка, а путь, идущий через моховидные волокна, наоборот, снимает это тормозное влияние.
Следовательно, деятельность всей нейрональной системы коры мозжечка сводится к торможению ядер, над которыми кора надстроена. Согласно гипотезе, высказанной Дж. Экклсом, большое количество тормозных нейронов в коре мозжечка предотвращает длительную циркуляцию возбуждения по нейронным цепям. Любой возбуждающий импульс, приходя в кору мозжечка, превращается в торможение за время порядка 100 мс. Так происходит как бы автоматическое стирание предшествующей информации, которое позволяет коре мозжечка участвовать в регуляции быстрых движений.
С точки зрения функций, которые выполняет мозжечок, его делят на три части: архиоцеребеллум (древний мозжечок), палеоцеребеллум (старый мозжечок) и неоцеребеллум (новый мозжечок).
Кора архиоцеребеллума (внутренняя часть – червь мозжечка) связана с ядром шатра, которое регулирует активность вестибулярных ядер. Поэтому архиоцеребеллум является вестибулярным регулятором. Повреждения древних структур мозжечка приводят к нарушению равновесия.
Функция палеоцеребеллума , или средней части коры – взаимная координация позы и целенаправленного движения, а также коррекция выполнения сравнительно медленных движений по механизму обратной связи – реализуется с участием пробковидных и шаровидных ядер. При повреждении структур старого мозжечка больным трудно стоять и ходить, особенно в темноте, при отсутствии зрительной коррекции.
Неоцеребеллум (латеральная часть коры мозжечка) совместно с зубчатым ядром играет важную роль в программировании сложных движений, выполнение которых идет без использования механизма обратных связей. В итоге возникает целенаправленное движение, выполняемое с большой скоростью, например, игра на фортепиано. При нарушении структур неоцеребеллума нарушаются сложные последовательности движений, они становятся аритмичны и замедлены.
Функции мозжечка изучались в клинике при его поражениях у человека, а также у животных путем удаления (экстирпации мозжечка) (Л. Лючиани, Л. А. Орбели). В результате выпадения функций мозжечка возникают двигательные расстройства (на стороне поражения), которые итальянский физиолог Л. Лючиани охарактеризовал знаменитой триадой А – астазия, атония и астения. Последующие исследователи добавили еще один симптом – атаксия (тетрада А).
Астазия – утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т. д., невозможность сохранения неподвижного положения, непрерывные качательные движения, дрожание головы, туловища и конечностей.
Атония – резкое падение и неправильное распределение тонуса мышц.
Астения – снижение силы мышечного сокращения, быстрая утомляемость мышц.
Атаксия – нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются также адиадохокинез, асинергия, пьяная шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз-вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Врожденных двигательных актов у человека не так уж много (например, сосание), большинство же движений он выучивает в течение жизни, и они становятся автоматическими (ходьба, письмо и т. д.). Когда нарушается функция мозжечка, движения становятся неточными, негармоничными, разбросанными, часто не достигают цели.
Дисметрия – несоответствие между интенсивностью мышечного сокращения и задачей выполняемого движения, что может выражаться в понижении или повышении интенсивности рефлекторных реакций. Например, человек, поднимаясь по лестнице, слишком высоко поднимает колени («петушиная походка»), или наоборот, спотыкается о каждую ступеньку.
Дизартрия – расстройство организации речевой моторики. При повреждении мозжечка речь больного становится растянутой, слова иногда произносятся как бы толчками (скандированная речь).
С течением времени двигательные расстройства сглаживаются. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации). Как показал Э. А. Асратян, компенсация функций происходит за счет коры головного мозга. Мозжечок участвует в регуляции движений, делая их плавными, точными, соразмерными.
Мозжечок оказывает влияние также на ряд вегетативных функций, например желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Долгое время мозжечок считался структурой, ответственной исключительно за координацию движений. Сегодня ученые все чаще говорят о его участии в процессах восприятия и когнитивной деятельности. Так, нейробиологи, изучающие когнитивные функции головного мозга, обнаружили, что у людей эта структура сохраняет высокую активность во время разнообразных форм деятельности, не связанных непосредственно с движениями.
 2. 3. 4 Средний мозг (mesencephalon).
Средний мозг расположен над мостом и представлен ножками мозга и четверохолмием. Ножки мозга состоят из основания и покрышки, между которыми находится черная субстанция, содержащая сильно пигментированные клетки. В покрышке мозга располагаются ядра блокового (IV пара) и глазодвигательного (III пара) нервов. Полость среднего мозга представлена узким каналом – сильвиевым водопроводом, который соединяет III и IV мозговые желудочки. Длина среднего мозга у взрослого человека около 2 см, вес – 26 г. В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются латерально и образуют сетчатку глаза, которая структурно и функционально представляет собой вынесенный на периферию нервный центр среднего мозга.
2. 3. 4 Средний мозг (mesencephalon).
Средний мозг расположен над мостом и представлен ножками мозга и четверохолмием. Ножки мозга состоят из основания и покрышки, между которыми находится черная субстанция, содержащая сильно пигментированные клетки. В покрышке мозга располагаются ядра блокового (IV пара) и глазодвигательного (III пара) нервов. Полость среднего мозга представлена узким каналом – сильвиевым водопроводом, который соединяет III и IV мозговые желудочки. Длина среднего мозга у взрослого человека около 2 см, вес – 26 г. В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются латерально и образуют сетчатку глаза, которая структурно и функционально представляет собой вынесенный на периферию нервный центр среднего мозга.
Наиболее крупными ядрами среднего мозга являются красные ядра, черная субстанция, ядра черепных (глазодвигательного и блокового) нервов и ядра ретикулярной формации. Через средний мозг проходят восходящие пути к таламусу, большим полушариям и мозжечку, и нисходящие пути к продолговатому и спинному мозгу.
В среднем мозге располагается большое количество нейронов ретикулярной формации. В четверохолмии выделяют верхнее и нижнее двухолмия (рис. 23).
Средний мозг выполняет ряд функций: проводниковую, двигательную и рефлекторную.
Проводниковая функция заключается в том, что через него проходят все восходящие пути к вышележащим отделам: таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция реализуется за счет ядра блокового нерва (n. trochlearis), ядер глазодвигательного нерва (п. oculomotorius), красного ядра (nucleus ruber), черного вещества (substantia nigra).
Красные ядра располагаются в покрышке (рис. 23.– В, 4). Будучи составной частью организующей движения экстрапирамидной системы, красные ядра получают нервные импульсы от моторной коры, подкорковых ядер, ядер мозжечка, черной субстанции среднего мозга и дают начало руброспинальному (красноядерно-спинномозговому) тракту, который как общий путь обеспечивает регуляцию тонуса скелетных мышц, снижая тонус мышц-разгибателей. Об этом свидетельствует классический опыт Ч. Шеррингтона с перерезкой стволовой части мозга. Если перерезку произвести на уровне переднего края задних холмов четверохолмия и таким образом отделить красное ядро от заднего мозга, то у кошки развивается децеребрационная ригидность . Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра, вышележащих структур и мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Со средним мозгом связано осуществление выпрямительных и статокинетических рефлексов. Выпрямительные рефлексы состоят из двух фаз: подъема головы и последующего подъема туловища. Первая фаза осуществляется вследствие рефлекторных влияний с рецепторов вестибулярного аппарата и кожи, вторая – с проприорецепторов мышц шеи и туловища. Статокинетические рефлексы направлены на возвращение тела в исходное положение при перемещении тела в пространстве, при вращении.
Другое функционально важное ядро среднего мозга – черная субстанция (Земмеринга) (рис. 23. – 5). Оно связано с лежащими в основании полушарий переднего мозга базальными ганглиями – полосатым телом и бледным шаром – и регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах за счет изменения их коллоидных свойств, что обеспечивает наименьшие затраты энергии. Регуляция этого процесса осуществляется клетками черного вещества.
Повреждение черной субстанции, вызывающее дегенерацию дофаминэргических путей к полосатому телу, связано с тяжелым неврологическим заболеванием – болезнью Паркинсона. Паркинсонизм проявляется в нарушении тонких содружественных движений, функции мимической мускулатуры и в появлении непроизвольных мышечных сокращений, или тремора. Этот болезненный синдром может быть снят при введении L-диоксифенилаланина – вещества, из которого синтезируется дофамин в организме.
Таким образом, восполняя дефицит медиатора, стало возможным купировать неврологическое заболевание и вместе с тем привести фактические доказательства роли черной субстанции среднего мозга в сенсомоторной координации движений.
Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние – слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными. В последнем случае они реагируют только на один признак раздражения, например смену света и темноты, направление движения светового источника и т. д. Основная функция бугров четверохолмия – организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные (верхнее двухолмие) или звуковые (нижнее двухолмие) сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию или к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы.
У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство. При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
2. 3. 5 Промежуточный мозг (diencephalon). Промежуточный мозг располагается под мозолистым телом и сводом, срастаясь по бокам с полушариями большого мозга. К нему относятся: таламус (зрительные бугры), гипоталамус (подбугорная область), эпиталамус (надбугорная область) и метаталамус (забугорная область). Полостью промежуточного мозга является III желудочек мозга.
Эпиталамус включает в себя железу внутренней секреции – эпифиз (шишковидное тело). В темноте она вырабатывает гормон мелатонин, который тормозит половое созревание, а также влияет на рост скелета.
Метаталамус представлен латеральными и медиальными коленчатыми телами. Латеральное , или наружное, коленчатое тело – это подкорковый центр зрения имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
 Медиальное коленчатое тело –
подкорковый, таламический центр слуха, в него поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферентные пути от медиальных коленчатых тел идут в височную долю коры большого мозга, достигая там первичной слуховой зоны. Медиальное коленчатое тело имеет четкую тонотопичность. Следовательно, уже на уровне промежуточного мозга обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Медиальное коленчатое тело –
подкорковый, таламический центр слуха, в него поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферентные пути от медиальных коленчатых тел идут в височную долю коры большого мозга, достигая там первичной слуховой зоны. Медиальное коленчатое тело имеет четкую тонотопичность. Следовательно, уже на уровне промежуточного мозга обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Таламус (thalamus, зрительный бугор) – парный орган яйцевидной формы (рис. 24) передняя часть которого заострена (передний бугорок), а задняя расширенная часть (подушка) нависает над коленчатыми телами. Медиальная поверхность таламуса обращена в полость III желудочка мозга.
Таламус – чувствительное ядро подкорки. Его называют «коллектором чувствительности», так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, исключая обонятельные.
В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприо- и интероцепторов на начинающиеся здесь таламокортикальные пути.
Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. В таламусе происходит сопоставление информации, получаемой по различным каналам, и оценка ее биологического значения. В зрительном бугре насчитывается около 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные. Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки – полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III – IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Нейроны неспецифических ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из ретикулярной формации ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния. Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом. Конвергенция сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
В клинике симптомами поражения зрительных бугров являются сильная головная боль, расстройства сна, нарушения чувствительности, как в сторону повышения, так и понижения, нарушения движений, их точности, соразмерности, возникновение насильственных непроизвольных движений.
Гипоталамус (hypothalamus, подбугорье) – структура промежуточного мозга, входящая в лимбическую систему, организующая эмоциональные, поведенческие, гомеостатические реакции организма (рис. 25).
Гипоталамус имеет большое число нервных связей с корой большого мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом.
 В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 2) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 5) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра.
В состав гипоталамуса входят серый бугор, воронка с нейрогипофизом и сосцевидные тела. Морфологически в нейронных структурах гипоталамуса можно выделить около 50 пар ядер, имеющих свою специфическую функцию. Топографически эти ядра можно объединить в 5 групп: 1) передняя группа, в состав которой входят супраоптическое, паравентрикулярные ядра; 2) задняя группа сформирована из медиальных и латеральных ядер сосцевидных тел и заднего гипоталамического ядра; 3) средняя группа состоит из нижнемедиального и верхнемедиального ядер; 4) преоптическая группа имеет выраженные связи с конечным мозгом и делится на медиальное и латеральное предоптические ядра; 5) наружная группа включает в себя латеральное гипоталамическое поле и серобугорные ядра.
Ядра гипоталамуса имеют мощное кровоснабжение, подтверждением чему служит тот факт, что ряд ядер гипоталамуса обладает изолированным дублирующим кровоснабжением из сосудов артериального круга большого мозга (виллизиев круг). На 1 мм 2 площади гипоталамуса приходится до 2600 капилляров, в то время как на той же площади V слоя предцентральной извилины (моторной коры) их 440, в гиппокампе – 350, в бледном шаре – 550, в затылочной доле коры большого мозга (зрительной коре) – 900. Капилляры гипоталамуса высокопроницаемы для крупномолекулярных белковых соединений, к которым относятся нуклеопротеиды, что объясняет высокую чувствительность гипоталамуса к нейровирусным инфекциям, интоксикациям, гуморальным сдвигам.
У человека гипоталамус окончательно созревает к 13 – 14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой большого мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К этим особенностям относятся чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреции пептидов, нейромедиаторов и др.
Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
Раздражение ядер передней группы сопровождается парасимпатическими эффектами. Раздражение ядер задней группы вызывает симпатические эффекты в работе органов. Стимуляция ядер средней группы приводит к снижению влияний симпатического отдела вегетативной нервной системы. Указанное распределение функций гипоталамуса не абсолютно. Все структуры гипоталамуса способны в разной степени вызывать симпатические и парасимпатические эффекты. Следовательно, между структурами гипоталамуса существуют функциональные взаимодополняющие, взаимокомпенсирующие отношения.
В целом за счет большого количества связей, полифункциональности структур гипоталамус выполняет интегрирующую функцию вегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций. Так, в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости. Все эти центры реализуют свои функции путем активации или торможения автономного (вегетативного) отдела нервной системы, эндокринной системы, структур ствола и переднего мозга.
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза. В гипоталамусе и гипофизе образуются также нейрорегуляторные пептиды – энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса и т. д.
Нейроны ядер передней группы гипоталамуса продуцируют вазопрессин, или антидиуретический гормон (АДГ), окситоцин и другие пептиды, которые по аксонам попадают в заднюю долю гипофиза – нейрогипофиз.
Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы , стимулирующие (либерины) и ингибирующие (статины) активность передней доли гипофиза – аденогипофиз. В нем образуются такие вещества, как соматотропный, тиреотропный и другие гормоны. Наличие такого набора пептидов в структурах гипоталамуса свидетельствует о присущей им нейросекреторной функции.
Нейроны гипоталамуса также обладают детектирующей функцией: реагируют на изменения температуры крови, электролитного состава и осмотического давления плазмы, количества и состав гормонов крови.
Исследования Дельгадо (Delgado) во время хирургических операций показали, что у человека раздражение определенных участков гипоталамуса вызывало эйфорию, эротические переживания. В клинике показано также, что патологические процессы в гипоталамусе могут сопровождаться ускорением полового созревания, нарушением менструального цикла, половой функции.
Раздражение передних отделов гипоталамуса может вызывать у животных пассивно-оборонительную реакцию, ярость страх, а раздражение заднего гипоталамуса вызывает активную агрессию. Кроме того, раздражение заднего гипоталамуса приводит к экзофтальму, расширению зрачков, повышению кровяного давления, сужению просвета артериальных сосудов, сокращениям желчного, мочевого пузырей. Могут возникать взрывы ярости с описанными симпатическими проявлениями. Уколы в области гипоталамуса вызывают глюкозурию, полиурию. В ряде случаев раздражение вызывало нарушение теплорегуляции: животные становились пойкилотермными, у них не возникало лихорадочное состояние.
Гипоталамус является также центром регуляции цикла бодрствование – сон. При этом задний гипоталамус активизирует бодрствование, стимуляция переднего вызывает сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон.
2. 3. 6 Конечный мозг (telencephalon). Конечный мозг является наиболее молодым в филогенетическом отношении. Он состоит из двух полушарий, каждое из которых представлено плащом, обонятельным мозгом и базальными или подкорковыми ганглиями (ядрами). Длина полушарий в среднем – 17 см, высота – 12 см. Полостью конечного мозга являются боковые желудочки, находящиеся в каждом из полушарий. Полушария большого мозга отделены друг от друга продольной щелью большого мозга и соединяются при помощи мозолистого тела, передней и задней спаек и спайки свода. Мозолистое тело состоит из поперечных волокон которые в латеральном направлении продолжаются в полушария, образуя лучистость мозолистого тела.
Обонятельный мозг представлен обонятельными луковицами, обонятельным бугорком, прозрачной перегородкой и прилежащими областями коры (препериформной, периамигдалярной и диагональной). Это самая меньшая часть конечного мозга, он обеспечивает функцию первого органа чувств, появившегося у живых существ, – функцию обоняния, и, кроме того, входит в состав лимбической системы. Повреждение структуры лимбической системы вызывает глубокое нарушение эмоций и памяти.
 Базальные ганглии
(ядра серого вещества) расположены в глубине больших полушарий. Они составляют примерно 3% от их объема. Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга (корой больших полушарий, таламусом, черной субстанцией, красным ядром, мозжечком, мотонейронами спинного мозга). К базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро
(рис. 26.–1) и заложенное в толще белого вещества чечевицеобразное ядро
. Двумя белыми пластинками оно подразделяется на наиболее крупную, лежащую латерально скорлупу
и бледный шар
(рис. 26.–2, 3). Хвостатое ядро и скорлупа объединяются под названием полосатого тела
, они связаны анатомически и характеризуются чередованием белого и серого вещества.
Базальные ганглии
(ядра серого вещества) расположены в глубине больших полушарий. Они составляют примерно 3% от их объема. Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга (корой больших полушарий, таламусом, черной субстанцией, красным ядром, мозжечком, мотонейронами спинного мозга). К базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро
(рис. 26.–1) и заложенное в толще белого вещества чечевицеобразное ядро
. Двумя белыми пластинками оно подразделяется на наиболее крупную, лежащую латерально скорлупу
и бледный шар
(рис. 26.–2, 3). Хвостатое ядро и скорлупа объединяются под названием полосатого тела
, они связаны анатомически и характеризуются чередованием белого и серого вещества.
Полосатое тело принимает участие в организации и регуляции движений и обеспечении перехода одного вида движения в другое. Стимуляция хвостатого ядра тормозит восприятие зрительной, слуховой и других видов сенсорной информации, угнетает активность коры, подкорки, безусловные рефлексы (пищевой, оборонительный и др.) и выработку условных рефлексов, приводит к наступлению сна. При поражении полосатого тела наблюдается ретроантероградная амнезия – выпадение памяти на события, предшествующие травме. Двустороннее повреждение полосатого тела побуждает к стремлению движения вперед, одностороннее – приводит к манежным движениям. С нарушением функций полосатого тела связывают заболевание нервной системы – хорею (усиливаются вспомогательные и мимические движения). Скорлупа обеспечивает организацию пищевого поведения. При ее поражении наблюдаются трофические нарушения кожи, а ее раздражение вызывает слюноотделение и изменение дыхания.
Функции бледного шара заключаются в провоцировании ориентировочной реакции, движения конечностей, пищевого поведения (жевание, глотание).
После разрушения бледного шара возникают скованность движений, обеднение мимики (маскообразное лицо), гиподинамия, эмоциональная тупость, тремор головы, конечностей при движении, монотонная речь. При повреждениях бледного шара могут появиться подергивания отдельных мышц лица и туловища, нарушается синергизм движения конечностей при ходьбе. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно.
Плащ у человека представлен корой , т. е. пластинкой серого вещества, отделенной от полости желудочков белым веществом , которое содержит огромное количество нервных волокон, подразделяемых на три группы.
1. Ассоциативные волокна , или пути, соединяют различные отделы коры большого мозга внутри одного полушария. Выделяют короткие или дугообразные ассоциативные волокна, связывающие две лежащие рядом извилины, и длинные – протягивающиеся из одной доли в другую, оставаясь в пределах одного полушария.
2. Комиссуральны е или спаечные волокна связывают кору обоих полушарий. Самой большой комиссурой головного мозга является мозолистое тело.
3. Проекционные пути связывают кору большого мозга с периферией. Существуют центробежные (эфферентные, двигательные) волокна, несущие нервные импульсы из коры на периферию, и центростремительные (афферентные, чувствительные) волокна, несущие импульсы с периферии в кору больших полушарий.
 Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Высшим отделом ЦНС является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций.
Кора большого мозга делится на древнюю (archicortex), старую (paleocortex) и новую (neocortex). Древняя кора наряду с другими функциями имеет отношение к обонянию и обеспечению взаимодействия систем мозга. Старая кора включает поясную извилину, гиппокамп. У новой коры наибольшее развитие величины, дифференциации функций отмечается у человека. Толщина новой коры колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине.
Кора полушарий покрыта бороздами и извилинами (рис. 27). Благодаря этому поверхность коры значительно увеличивается. Различают самые глубокие первичные борозды, которые делят полушария на доли. Боковая борозда (Сильвиева) отделяет лобную долю от височной, центральная борозда (Роландова) – лобную от теменной. Теменно-затылочная борозда располагается на медиальной поверхности полушария и разделяет теменную и затылочную доли, на верхнелатеральной поверхности явная граница между этими долями отсутствует. На медиальной поверхности располагается поясная борозда, переходящая в гиппокампову борозду, которые ограничивают обонятельный мозг от остальных долей.
В глубине боковой борозды (рис. 28) располагается островковая доля . Она окружена с трех сторон круговой бороздой, ее поверхность изрезана бороздами и извилинами. Функционально островок связан с обонятельным мозгом.
 Вторичные борозды менее глубокие, они делят доли на извилины и располагаются снаружи от одноименных извилин. Третичные (безимянные) борозды придают извилинам индивидуальную форму, увеличивают площадь их коры.
Вторичные борозды менее глубокие, они делят доли на извилины и располагаются снаружи от одноименных извилин. Третичные (безимянные) борозды придают извилинам индивидуальную форму, увеличивают площадь их коры.
Функции отдельных зон новой коры определяются особенностями ее структурно-функциональной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов.
Особенности структурнофункцио- нальной организации коры большого мозга обусловлены тем, что в эволюции происходила кортикализация функций, т. е. передача коре большого мозга функций нижележащих структур мозга. Однако эта передача не означает, что кора берет на себя выполнение функций других структур. Ее роль сводится к коррекции возможных нарушений функций взаимодействующих с ней систем, более совершенного, с учетом индивидуального опыта, анализа сигналов и организации оптимальной реакции на эти сигналы, формирование в своих и в других заинтересованных структурах мозга памятных следов о сигнале, его характеристиках, значении и характере реакции на него. В дальнейшем, по мере автоматизации реакция начинает выполняться подкорковыми структурами.
Цитоархитектоника коры головного мозга. Общая площадь коры большого мозга человека около 2200 см 2 , число нейронов коры превышает 10 млрд. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков; аксон пирамидного нейрона, как правило, идет через белое вещество в другие зоны коры или в структуры ЦНС.
Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аскон, обеспечивающий связи нейронов в пределах самой коры большого мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
Кора большого мозга имеет преимущественно шестислойное строение (рис. 29).
 Слой I – верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой I – верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры большого мозга.
Слой II – наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре большого мозга, т. е. имеющих отношение к памяти.
Слой III – наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со II слоем обеспечивают корко-корковые связи различных извилин мозга.
Слой IV – внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокортикальные пути, т. е. пути, начинающиеся от рецепторов анализаторов.
Слой V – внутренний пирамидный (ганглиозный), слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг. В двигательной зоне в этом слое находятся гигантские пирамидные клетки, открытые Бецом (клетки Беца).
Слой VI – слой полиморфных клеток, большинство нейронов этого слоя образуют кортико-таламические пути.
Клеточный состав коры по разнообразию морфологии, функции, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека 53 цитоархитектонических поля (поля Бродмана). Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе.
У человека и высших млекопитающих различают, наряду с первичными, вторичные и третичные корковые поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов. Для всех анализаторов характерен соматотопический принцип организации
Особенностью корковых полей является экранный принцип их функционирования. Этот принцип заключается в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их коллатералями и связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так, одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1 мм. Это значит, что один аксон распределяет свое действие на более чем 5 000 нейронов.
Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам III – V слоев коры. От звездчатых клеток IV слоя сигнал идет к пирамидным нейронам III слоя, а отсюда по ассоциативным волокнам – к другим полям, областям коры большого мозга. Звездчатые клетки поля 3 переключают сигналы, идущие в кору, на пирамидные нейроны V слоя, отсюда обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые колонки – функциональные единицы коры, организованные в вертикальном направлении. Доказательством этого служит следующее: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.
Диаметр колонки около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в организации той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних.
Как уже упоминалось, разные области коры большого мозга имеют разные поля, определяющиеся по характеру и количеству нейронов, толщине слоев и т. д. Наличие структурно различных полей предполагает и разное их функциональное предназначение (рис. 30). Действительно, в коре большого мозга выделяют сенсорные, моторные и ассоциативные области.
Среди всех систем организма ЦНС занимает особое место. Головной мозг регулирует все функции, которыми наделён человек. Благодаря ему осуществляется взаимосвязь между работой органов и систем. Без мозговой регуляции человек не был бы жизнеспособным существом. Благодаря скоординированной деятельности ЦНС мы двигаемся, говорим, думаем и чувствуем внешние раздражители. Головной мозг имеет сложное строение, каждая из его составляющих отвечает за определённую функцию. Тем не менее все его структуры обеспечивают работу нашего организма лишь в совокупности. Особо важными образованиями, входящими в состав ЦНС, являются и варолиев мост. Именно они содержат основные жизненно важные центры (сосудистый, дыхательный, кашлевой, слёзоотделительный), а также дают начало большинству черепных нервов.
Строение головного мозга
Структурной единицей ЦНС является нейрон. Именно эта клетка отвечает за приём, переработку и хранение информации. Весь представляет собой скопления нейронов и их отростков - аксонов и дендритов. Они обеспечивают передачу сигналов, поступающих в ЦНС и обратно, к органам. Головной мозг состоит из серого и Первое образуют сами нейроны, второе - их аксоны. Основными структурами головного мозга являются и правое), мозжечок и ствол. Первые отвечают за умственные способности человека, его память, мышление, воображение. Мозжечок необходим для координации движений, в частности, он обеспечивает возможность ровно стоять, ходить, брать предметы. Под ним располагается варолиев мост. Он является связующим звеном между продолговатым мозгом и мозжечком.
Варолиев мост: строение и функции
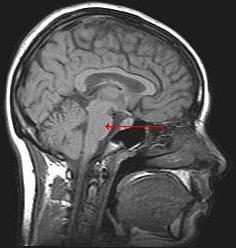
Мост является одной из частей заднего мозга. Его длина колеблется в пределах от 2,4 до 2,6 см. Варолиев мост имеет массу около 7 г. Структуры, которые с ним граничат, - это продолговатый и поперечная борозда. Основными составляющими варолиева моста являются верхние и средние ножки мозжечка, которые являются крупными проводящими путями. Спереди находится базилярная борозда, содержащая в себе артерии, питающие головной мозг, а рядом расположено место выхода С задней стороны варолиев мост образует верхнюю часть ромбовидной ямки, в которой заложены 6 и часть 7 черепно-мозговых нервов. В верхней части моста залегает больше всего ядер (5, 6, 7, 8). В основании моста имеются проводящие нисходящие пути: кортикоспинальный, бульбарный и мостовой тракты.
Основные функции этого органа:
- Проводниковая - по его путям проходят нервные импульсы к коре больших полушарий и к спинному мозгу.
- Сенсорная функция - обеспечивается благодаря преддверно-улитковому и тройничному нервам. В ядрах 8-й пары черепно-мозговых нервов происходит обработка информации о вестибулярных раздражениях.
- Двигательная - обеспечивает сокращение всех Это происходит благодаря ядрам тройничного нерва. Помимо этого, его чувствительная часть получает информацию от рецепторов слизистой оболочки рта, глазного яблока, части головы и зубов. Эти сигналы по волокнам моста направляются в кору больших полушарий.
- Интегративная функция обеспечивает взаимосвязь между передним и задним мозгом.
- Рефлексы головного мозга.

Ретикулярная формация моста
Ретикулярная формация - это разветвлённая сеть, расположенная в головном мозге и состоящая из нервных клеток и ядер. Она имеется практически у всех образований ЦНС и плавно переходит из одного отдела в другой. Ретикулярная формация варолиева моста расположена между продолговатым и средним мозгом. Её длинные отростки - аксоны, образуют белое вещество и переходят в мозжечок. Кроме того, по волокнам нервных клеток моста сигналы могут переноситься из головного в спиной. Помимо этого, ретикулярная формация передаёт сигналы в кору больших полушарий, благодаря чему происходит пробуждение или сон человека. Ядра, расположенные в этой части моста, относятся к центру дыхания, расположенному в продолговатом мозге.
Рефлекторная функция моста
Способность центральной нервной системы отвечать на внешние раздражители называется рефлексом. Примером может служить появление слюнотечения при виде пищи, желание спать при звуке успокаивающей музыки и т. д. Рефлексы головного мозга могут быть условными и безусловными. Первые человек приобретает в процессе жизни, их можно нарабатывать или корректировать в зависимости от нашего желания. Вторые не поддаются сознанию, они заложены с рождением, и изменить их невозможно. К ним относится жевательный, глотательный, хватательный и другие рефлексы.

Как мост влияет на возникновение рефлексов
Благодаря тому что варолиев мост является составной частью четверохолмия, он имеет отношение к развитию слухового и статистического рефлекса. Благодаря последнему мы способны удерживать тело в определённой позе. Кроме того, взаимодействуя со средним мозгом, он замыкает значительную часть мышечных рефлексов.
166 ..Мост (анатомия мозга человека)
Мост , pons, является частью мозгового ствола длиной 25 мм, которая расположена между продолговатым и средним мозгом. Его вентральная поверхность образована выпуклостью белого цвета, снаружи состоящей из поперечно расположенных волокон. Дорсальная поверхность моста составляет верхнюю часть дна IV желудочка - ромбовидной ямки, образуя ее верхний треугольник. Эта часть ромбовидной ямки ограничена верхними мозжечковыми ножками. Поперечные волокна передней поверхности образуют средние мозжечковые ножки, которые погружаются в толщу полушарий мозжечка. Границей между мостом и средней мозжечковой ножкой служит linea trigeminofacialis, проходящая между корешками тройничного и лицевого нервов. Посередине вентральной поверхности моста расположена основная борозда, sulcus basilaris. В боковом отделе вентральной поверхности моста, ближе к его переднему краю, находятся корешки тройничного нерва. В области мосто-мозжечкового угла (образован продолговатым мозгом, мостом и мозжечком) расположены корешки лицевого, промежуточного и преддверно-улиткового нервов, а ближе к средней линии, между задним краем моста и пирамидами, - корешки отводящих нервов.
На поперечных срезах моста различают крупную вентральную часть, pars ventralis pontis, и меньшую - дорсальную, pars dorsalis pontis, границей между которыми является пучок поперечных волокон - трапециевидное тело, corpus trapezoideum. Вентральная и дорсальная части моста образованы серым и белым веществом. Однако вентральная часть состоит преимущественно из белого вещества.
Серое вещество вентральной части слагается из многочисленных собственных ядер моста, nuclei pontis. В этих ядрах заканчиваются корково-мостовые пути, tractus corticopontinus, и коллатерали от пирамидных путей. Волокна клеток ядер моста образуют поперечные волокна моста, fibrae pontis transversae, которые, в основном, переходя на противоположную сторону, составляют средние ножки мозжечка и заканчиваются в клетках коры его полушарий. Поперечные волокна моста в вентральной части его нижнего отдела образуют поверхностный и глубокий слои, между которыми проходят пучки пирамидного пути. В верхних отделах моста глубокий слой его поперечных волокон увеличивается в объеме и появляется третий слой, разделяющий пирамидные пути на более мелкие пучки.
Серое вещество дорсальной части моста состоит из центрально расположенного сетчатого образования, ядер черепных нервов и переключательных ядер. Сетчатое образование моста является непосредственным продолжением одноименного образования продолговатого мозга.
В мосту расположены ядра следующих черепных нервов: отводящего, лицевого, тройничного, преддверно-улиткового.
Ядро отводящего нерва, nucleus n. abducentis, образовано крупными двигательными клетками. Оно расположено возле средней линии дна IV желудочка и имеет длину около 3 мм. Отростки клеток ядра выходят через толщу моста между его задним краем и пирамидой в виде корешка отводящего нерва.
Ядро лицевого нерва, nucleus n. facialis, длиной от 2 до 5,6 мм, образовано двигательными клетками. Оно расположено в сетчатом образовании дорсальной части моста. Отростки клеток этого ядра образуют внутримозговой отдел корешка лицевого нерва, который в толще моста имеет сложный ход. Корешок из сетчатого образования проходит по дну IV желудочка, образует колено, genu n. facialis, окружающее ядро отводящего нерва и далее идет вперед через толщу моста в область мостомозжечкового угла. Волокна лицевого нерва распределяются в мимической мускулатуре и осуществляют ее двигательную иннервацию. Несколько кзади от ядра лицевого нерва в сетчатом образовании моста находится верхнее слюноотделительное ядро, nucleus salivatorius superior, которое является секреторным вегетативным центром иннервации подъязычной и подчелюстной слюнных желез и слезной железы. Чувствительным ядром этого нерва является ядро одиночного пути, nucleus tractus solitarii, где оканчиваются центральные отростки узла промежуточного нерва - узла колена, gangl. geniculi, расположенного в канале лицевого нерва пирамиды височной кости.
В среднем и верхнем отделах дорсальной части моста расположены чувствительное и двигательное ядро тройничного нерва. Двигательное ядро тройничного нерва, nucleus motorius n. trigemini, образовано крупными двигательными клетками и имеет длину около 4 мм. Отростки клеток двигательного ядра образуют двигательный корешок тройничного нерва и распределяются в жевательной мускулатуре, осуществляя ее двигательную иннервацию. В составе двигательного корешка тройничного нерва проходят также волокна от ядра тройничного нерва, расположенного в среднем мозге, латеральнее водопровода большого мозга. Верхнее чувствительное ядро тройничного нерва, nucleus sensorius n. trigemini superior, лежит кнаружи от двигательного ядра. Оно меньше, чем ядро спинномозгового тракта тройничного нерва. В клетках этого ядра оканчиваются центральные отростки чувствительных клеток тройничного узла. Эти отростки образуют чувствительный корешок тройничного нерва, который через толщу базальной части моста подходит к верхнему чувствительному ядру. Здесь часть волокон корешка заканчивается, а остальные волокна проходят к ядрам спинномозгового и среднемозгового путей Тройничного нерва. Отростки клеток верхнего чувствительного ядра спинномозгового тракта (вторые нейроциты) переходят на противоположную сторону и в составе медиальной петли вступают в зрительный бугор.
К переключательным ядрам дорсальной части моста относятся верхние оливы, ядра трапециевидного тела и боковой петли. Во всех этих ядрах происходит переключение импульсов слухового пути.
Верхняя олива, oliva superior, расположена в латеральных отделах трапециевидного тела, которое образуют преимущественно отростки клеток вентрального ядра n. vestibulocochlearis. Между волокнами трапециевидного тела находятся скопления серого вещества - вентральное и дорсальное ядра трапециевидного тела, nuclei ventralis et dorsalis corporis trapezoidei. Большая часть волокон, которые возникают в вентральном ядре n. vestibulocochlearis, переходят на противоположную сторону и заканчиваются в верхней оливе и ядрах трапециевидного тела. Меньшая часть этих волокон заканчивается в соответствующих ядрах своей стороны. Отростки клеток верхней оливы образуют так называемую боковую петлю, lemniscus lateralis, среди волокон которой находится ядро боковой петли, nucleus lemnisci lateralis. Боковая петля представляет собой значительный по толщине пучок волокон; в ее состав входят отростки клеток дорсального ядра n. vestibulocochlearis, а также отростки клеток ядра трапециевидного тела и ядра боковой петли. Боковая петля заканчивается в первичных слуховых центрах - нижних буграх четверохолмия и медиальном коленчатом теле.
Белое вещество моста состоит из эндогенных и экзогенных волокон. Короткие эндогенные волокна соединяют отдельные ядра моста и не выходят за его пределы. Так, установлены нервные связи между ядрами лицевого и тройничного нервов, посредством которых замыкаются рефлекторные дуги при различных раздражениях кожи лица с ответной реакцией мимических мышц. Длинные эндогенные волокна возникают в ядрах моста и заканчиваются в других отделах центральной нервной системы.
К этой группе относятся волокна, идущие от собственных ядер моста к мозжечку - поперечные волокна моста, fibrae pontis transversae, волокна латеральной петли, пучки волокон от некоторых черепных нервов. Поперечные волокна моста образуют среднюю мозжечковую ножку, посредством которой осуществляется влияние коры полушарий головного мозга на деятельность мозжечка. 166 ..
 (1 оценок, в среднем: 5,00 из 5)
(1 оценок, в среднем: 5,00 из 5)

